Original Editor – Joanne Garvey and Naomi O’Reilly
Top Contributors – Joanne Garvey, Lucinda hampton, Rachael Lowe, Naomi O’Reilly and Tarina van der Stockt
Introduction
Neurophysiologi er den gren af fysiologien, der beskæftiger sig med nervesystemets funktioner. Dvs. studiet af de funktionelle egenskaber af neuroner, glia og netværk.
- Historisk set har den været domineret af elektrofysiologi – den elektriske registrering af neuronale hændelser lige fra det molære (elektroencefalogrammet, EEG) til det cellulære (intracellulær registrering af egenskaberne hos enkelte neuroner).
- Da neuronet er en elektrokemisk maskine, er det umuligt at adskille elektriske hændelser fra de biokemiske og molekylære processer, der fremkalder dem.
- Neurofysiologer anvender i dag teknikker fra kemi (calciumafbildning), fysik (funktionel magnetisk resonansafbildning, fMRI) og molekylærbiologi (stedbestemte mutationer) til at studere hjernens funktion.
Nedenfor vil du lære alt om
- Ionkanaler
- Hvilemembran og aktionspotentiale
- Neuromuskulær forgrening / synapser
- Nervekonduktion
- Neurotransmittere, Receptorer og veje
Ionkanaler
En ionkanal er et proteinmakromolekyle, der krydser membranens bredde og tillader molekyler at passere igennem. Ionerne bevæger sig i en retning, der er bestemt af den elektrokemiske gradient på tværs af membranen.
- Ioner har en tendens til at strømme fra et område med høj koncentration til et område med lav koncentration.
- I tilstedeværelsen af en spændingsgradient kan der ikke forekomme nogen strøm af ioner på trods af ulige koncentrationer.
- Ionkanaler kan være åbne eller lukkede.
- Åbning sker ved at ændre spændingen over membranen eller ved at binde et kemisk stof til en receptor.
- Den vigtigste rolle er, at de giver neuronet elektrisk excitabilitet.
- Findes i alle dele af neuronet og i mindre grad i neurogliacellerne.
Typer af kanaler
- Spændingsaktiveret
- Kemisk aktiveret
- Mekanisk strækning/tryk.
Fundamentale egenskaber ved en ionkanal
- Den består af en række proteinunderenheder, der sidder på tværs af membranen, og som tillader ioner at krydse fra den ene side til den anden. (Transmembranporen)
- Kanalen skal kunne bevæge sig fra åben til lukket tilstand og tilbage igen.
- Den skal kunne åbne sig som reaktion på de relevante stimuli.
Nogle kanaler reagerer på kemiske stimuli (især ved synapsen). Disse kanaler har specifikke receptorer for det pågældende kemikalie, der fører til kanalåbning.
Restmembran og aktionspotentiale
- I hviletilstand er den neuronale cellemembran temmelig uigennemtrængelig for ioner. Dette er afgørende for dannelsen af hvilemembranpotentialet.
- Den vigtigste intracellulære ion er kalium (det er natrium i den ekstracellulære væske).
- Den naturlige strøm af ioner ved hjælp af deres koncentrationsgradienter er, at K+ forlader cellen, og Na+ kommer ind.
- Denne bevægelse af ioner ud af cellen fører til et negativt membranpotentiale – Hyperpolarisering.
- Det modsatte er tilfældet ved en relativ tilstrømning af ioner – Hypopolarisering.

Den hvilende membran er relativt uigennemtrængelig for Na+-ioner, mens den forbliver gennemtrængelig for K+-ioner. Så når membranen er i hvile, vil der være en tendens til udstrømning af K+-ioner ud af cellen, ned ad dens koncentrationsgradient, hvorved overskydende negativ ladning efterlades. Dette fortsætter, indtil den kemiske koncentrationsgradient, der driver K+ ud af cellen, er præcis på det punkt, hvor den opvejes af den elektriske potentialforskel, der genereres af denne udstrømning. (membranpotentialet) K+ trækkes derefter tilbage ind i cellen.
Denne stationære tilstand i membranen kaldes ligevægtspotentialet.
Ligningen til udledning af ligevægtspotentialet kaldes Nernst-ligningen.
Der er faktisk en lidt uventet øget positiv ladning i axoner på grund af en lille permeabilitet for Na+-ioner i membranen i hviletilstand. Dette opvejes af den ATP-afhængige Na+/K+-udvekslingspumpe. Jeg pumper 3 Na+-ioner ud for hver 3K+-ioner, der bringes ind i cellen. Den yder dog kun et meget lille bidrag til cellens membranpotentiale i hvile.
Aktionspotentiale
Defineret som en “enkelt elektrisk impuls, der passerer ned gennem et axon”.
Det er alt eller intet i sin virkning. Det betyder, at når tærskelintensiteten for stimulus er nået, vil der blive genereret et aktionspotentiale.
Information i nervesystemet kodes og fortolkes af fyringsfrekvensen og ikke af størrelsen af aktionspotentialet.
Tærskel stimulusintensitet: Den værdi, ved hvilken nettostrømmen indad (bestemt af Na+-ioner) er lige større end nettostrømmen udad (båret af K+-ioner). Den er normalt -55mV (kritisk fyringstærskel)
Appet opstår lettest ved axonhullet, fordi det er her, der er en større tæthed af Na+ ionkanaler. Det er derfor, at det er her, at AP indledes i neuronet.
Hvis tærskelværdien ikke nås, vil aktionspotentialet ikke blive genereret, og transmissionen af signalet afsluttes på dette punkt.
Hændelsesforløb for at et aktionspotentiale kan genereres
- Depoloriserende spænding aktiverer de spændingsfølsomme Na+-ionkanaler i neuronmembranen: Na+-ioner strømmer ned ad den elektrokemiske gradient. Membranen depolariseres yderligere. Yderligere Na+-kanaler åbnes i en Positiv feedback-loop. Når der er en større indadgående strøm af natriumioner i forhold til udstrømningen af K+ -ioner, åbnes alle Na+ -kanaler hurtigt. Herved depolariseres membranen i retning af ligevægtspotentialet for Na+ (+55 mV). Der opstår en spike af AP, men det lykkes ikke at nå ligevægtspotentialet for Na+ på grund af det stigende K+udstrømningspotentiale.
- Når Na+-kanalerne bliver mindre aktive, falder AP. Denne inaktivering er spændingsafhængig. I den faldende fase er K+-strømmen vigtig, da den fører til en kort periode med membranhyperpolarisering, inden den deaktiveres.
- Membranpotentialet vender tilbage til hviletilstand.
Neuromuskulært kryds / synapser
Synapse
En synapse er krydset mellem to neuroner. Den kemiske synapse er den fremherskende synapse, der findes i nervesystemet, men elektriske synapser findes dog i hjertemuskler og gliaceller.
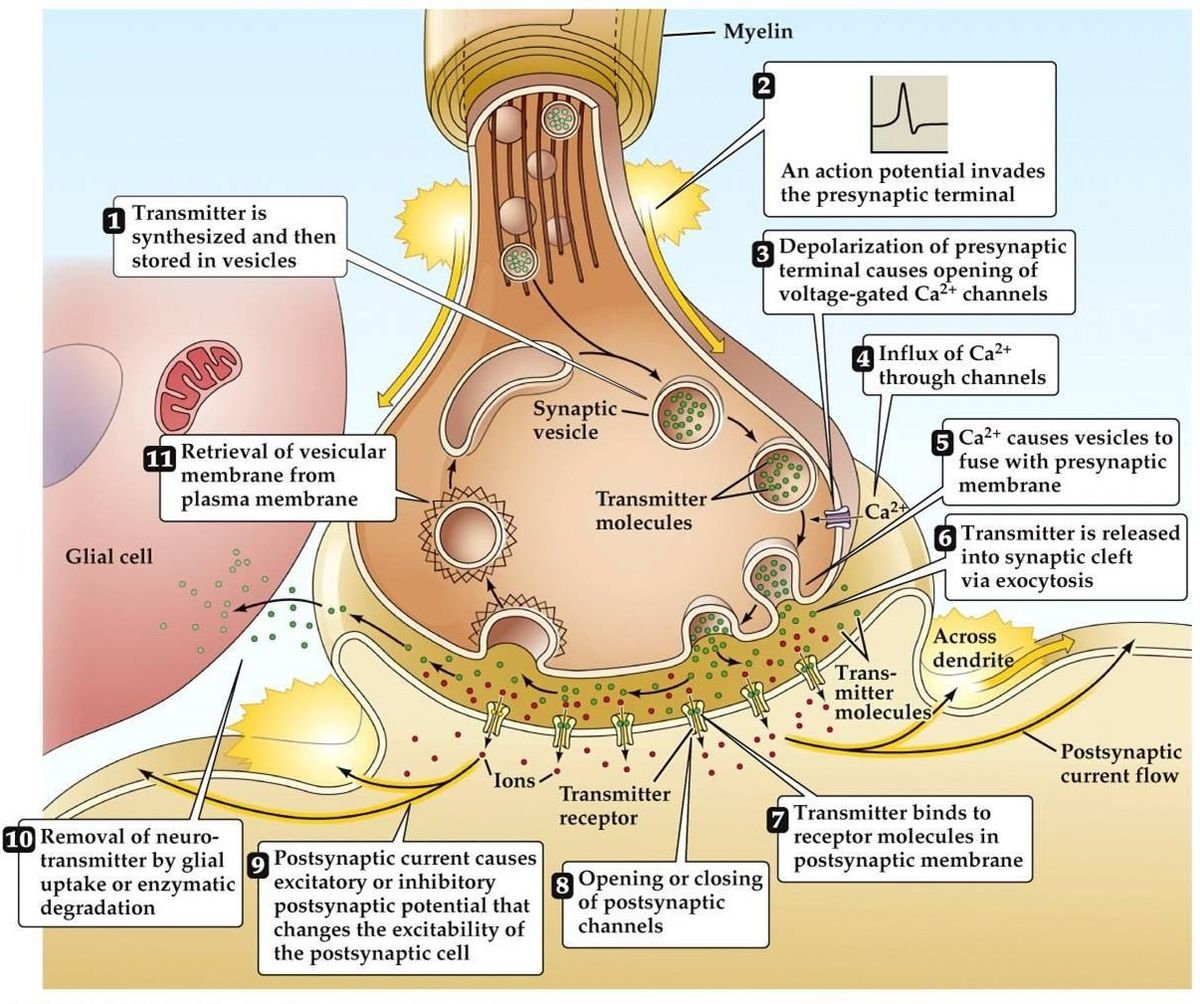
Synaptisk transmission
- Aktionspotentialet ankommer, fører til depolarisering af den præsynaptiske terminal. Spændingsafhængige Ca2+-kanaler åbnes i de aktive zoner af terminalen. Dette fører til tilstrømning af Ca2+.
- Influx af Ca2+ fører til fosforylering og ændring af mængden af presynaptiske calciumbindende proteiner. Dette frigør vesiklen fra dens præsynaptiske actinnetværk, Den binder sig derefter til den præsynaptiske membran.
- Fusionen af vesiklen til membranen fører til dannelsen af en lille kanal, som hurtigt udvider sig og frigiver sit indhold i den synaptiske kløft. Vesikelmembranen genanvendes ved endocytose.
- Den frigivne neurotransmitter diffunderer gennem den synaptiske kløft med det formål at binde sig til den postsynaptiske receptor.
- Aktivering af den postsynaptiske receptor fører til en ændring i det postsynaptiske membranpotentiale.
- Der er alternative teorier, der tyder på, at neurotransmitteren leveres via molekyler eller membrankanaler snarere end via vesikler.
Nerveledning
Aktionspotentialeudbredelse opnås ved lokal strømafspredning.
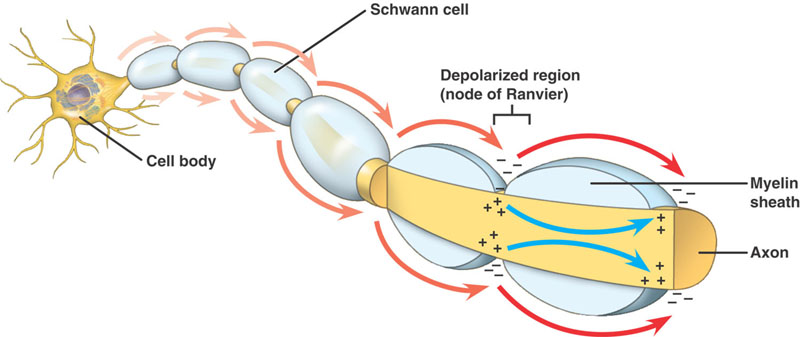
Nerven er isoleret med myelin, hvis dens størrelse er over en vis diameter, med Ranvier-knuder med forskellige intervaller langs dens længde.
Umyeliniserede axoner
Aktionspotentialet fører til depolarisering af membranen umiddelbart foran (og bagved). Membranen er i en refraktær tilstand, så aktionspotentialet ledes kun i 1 retning. Dette kan lade sig gøre i små axoner, men spredningen af strømmen er langsom.
Myeliniserede axoner
Samme hændelsesforløb som i de umyeliniserede axoner. Der er dog en væsentlig forskel. Det fremadskridende aktionspotentiale støder på en struktur med høj modstand og lav kapacitet – myelin, der er viklet rundt om axonet. Den depolariserende strøm passerer langs axoplasmaet, indtil den når Ranvier-knuden med lav modstand og dens store antal Na+-kanaler. Handlingspotentialet genereres derefter på dette sted. Aktionspotentialet ledes fra knude til knude. Dette kaldes Saltatory Conduction.
Fordele ved myelinisering
- Giver mulighed for hurtig ledelse af aktionspotentialet
- Minimerer de metaboliske krav til cellen.
- Gør NS’s pakningskapacitet større, så der kan proppes flere fibre ind i 1 nerve.
De fleste nerver større end 1 mikron er myeliniserede.
Forstyrrelser i ledningen skyldes normalt demyeliniseringspatologier. De omfatter Guillan Barre-syndromet og multipel sklerose.
Postsynaptisk integration:
Hvert centralt neuron modtager mange hundrede synapser. Hvert af disse input integreres derefter til et svar af det pågældende neuron. Dette indebærer behandling af alle de input, der ankommer på et hvilket som helst tidspunkt. Dette kaldes rumlig summation. Behandling af input over en bestemt tidsperiode kaldes tidsmæssig summation. Der vil være en blanding af excitatoriske og inhiberende synapser, der virker på dette punkt.
Ekscitatoriske postsynaptiske potentialer
- Binding af neurotransmitter fører til åbning af ionkanaler.
- Der sker kationindstrømning i postsynaptisk proces
- Depolarisering af membranen finder sted.
- EPSP’er depolariseringer registreret i den postsynaptiske celle til en bestemt excitatorisk stimulus.
- Depolariseringer assoc. med EPSP’er kan fortsætte med at udløse AP’er, hvis der sker summation i synapsen.
Hæmmende postsynaptiske potentialer
- Ionkanaler tillader postsynaptisk aniontilstrømning, når de er åbne.
- Hyperpolariseringer af membranen opstår på grund af indstrømning af CL- og udstrømning af K+
- Vigtigt for modulering af neuronernes respons på excitatorisk input ved synapsen.
- Findes på strategisk vigtige steder i neuronen som f.eks. den proximale dendrit og soma.
- De har stor indflydelse på det dendritiske træ
- Nogle neuroner giver deres egen hæmmende indflydelse ved at have axonkollateraler og deres egne hæmmende interneuroner. Dette betegnes som feedbackhæmning. F.eks. motorneuroner og Renshaw-celler i rygmarven.
Neurotransmittere, receptorer og veje
Neurotransmitteren frigives ved synapsen og arbejder sammen med et specifikt protein i den postsynaptiske membran, kaldet receptoren. I nogle synapser kan det også forekomme, at neurotransmitteren interagerer med en præsynaptisk autoreceptor. PSA fungerer som en regulator for mængden af den frigivne transmitter.
Receptorer er normalt specifikke for en bestemt neurotransmitter. Der findes flere typer af receptorer. Samtidig frigivne neurotransmittere kan regulere bindingen af en anden transmitter.
Receptorer for specifikke neurotransmittere kan enten være koblet direkte til ionkanaler eller til et membranenzym, hvor neurotransmitterens binding til receptoren enten åbner en ionkanal via en intracellulær enzymkaskade eller indirekte modulerer sandsynligheden for, at andre ionkanaler åbnes som reaktion på spændingsændringer. (neuromodulation)
En aktiveret receptor kan først vende tilbage til sin hviletilstand, når neurotransmitteren er fjernet ved processen med enzymatisk hydrolyse (optagelse) i den præsynaptiske nerve terminal eller i de nærliggende gliaceller.
Desensibilisering/nedregulering: Et fald i receptorens affinitet for transmitteren på kort sigt, på lang sigt fører dette til et fald i antallet af receptorer.
Supersensitivitet/opregulering: En øget affinitet af receptoren for transmitteren på kort sigt kan føre til en stigning i antallet af kanaler på lang sigt.
Receptorer:
- Ionotropisk:
- Ionotropisk: N-methyl-D-aspartat(NMDA)
- Non-NMDA.
- Metabotropiske : G-protein-associerede glutamatreceptorer, der reagerer ved at igangsætte intracellulære biokemiske hændelser og modulere den synaptiske transmission.
Der findes et stort antal neurotransmittere:
- Excitatoriske aminosyrer: Disse er de vigtigste excitatoriske NT’er i CNS. Den vigtigste NT i denne gruppe er glutamat, som virker på en række receptorer.
- Inhiberende aminosyrer: Dette er de vigtigste hæmmende neurotransmittere, hvoraf den vigtigste er GABA, der findes i hele CNS. Glycin, findes hovedsagelig i rygmarven.
- Monoaminer: Findes i små grupper af neuroner i hjernestammen, der projicerer bredt ud i CNS. Findes i det autonome NS. De binder sig til et væld af receptorer.
- Acetylcholin: Udbredt i hele nervesystemet, herunder i det neuromuskulære knudepunkt og det autonome NS.
- Neuropeptider: Findes over hele NS. Ofte frigives de samtidig med andre NT’er.
Dertil kommer, at der er fundet over 50 neuroaktive peptider, og der bliver jævnligt opdaget nye. Mange af disse “frigives samtidig” sammen med en småmolekylær transmitter. Ikke desto mindre er et peptid i nogle tilfælde den primære transmitter i en synapse. β-endorfin er et relativt velkendt eksempel på en peptid-neurotransmitter, fordi den indgår i meget specifikke interaktioner med opioidreceptorer i centralnervesystemet.
Enkle ioner (såsom synaptisk frigivet zink) betragtes også af nogle som neurotransmittere, og det samme gælder nogle gasformige molekyler såsom nitrogenoxid (NO), kulilte (CO) og svovlbrinte (H2S). Gasserne produceres i det neurale cytoplasma og diffunderes straks gennem cellemembranen til den ekstracellulære væske og til nærliggende celler for at stimulere produktionen af sekundære budbringere. Opløselige gasneurotransmittere er vanskelige at undersøge, da de virker hurtigt og straks nedbrydes og kun eksisterer i få sekunder.
Den mest udbredte transmitter er glutamat, som er excitatorisk på langt over 90 % af synapserne i den menneskelige hjerne. Den næstmest udbredte er gamma-aminosmørsyre eller GABA, som er hæmmende på mere end 90 % af de synapser, der ikke bruger glutamat. Selv om andre transmittere anvendes i færre synapser, kan de være meget vigtige funktionelt set: Langt de fleste psykoaktive stoffer udøver deres virkning ved at ændre virkningen af nogle neurotransmittersystemer, der ofte virker gennem andre transmittere end glutamat eller GABA. Afhængighedsskabende stoffer som f.eks. kokain og amfetamin udøver deres virkninger primært på dopaminsystemet. De vanedannende opiater udøver deres virkninger primært som funktionelle analoger af opioide peptider, som igen regulerer dopaminniveauerne.