Editorial original – Joanne Garvey y Naomi O’Reilly
Contribuidores principales – Joanne Garvey, Lucinda hampton, Rachael Lowe, Naomi O’Reilly y Tarina van der Stockt
- Introducción
- Canales iónicos
- Tipos de canales
- Propiedades fundamentales de un canal iónico
- Membrana en reposo y potencial de acción
- Potencial de acción
- Unión Neuromuscular / Sinapsis
- Sinapsis
- Transmisión sináptica
- Conducción nerviosa
- Axones no mielinizados
- Axones mielinizados
- Neurotransmisores, receptores y vías
Introducción
La neurofisiología es la rama de la fisiología que se ocupa de las funciones del sistema nervioso. Es decir, el estudio de las propiedades funcionales de las neuronas, la glía y las redes.
- Históricamente ha estado dominada por la electrofisiología-el registro eléctrico de los eventos neuronales que van desde lo molar (el electroencefalograma, EEG) a lo celular (registro intracelular de las propiedades de las neuronas individuales).
- Como la neurona es una máquina electroquímica, es imposible separar los eventos eléctricos de los procesos bioquímicos y moleculares que los provocan.
- Los neurofisiólogos utilizan hoy en día técnicas de química (imágenes de calcio), física (imágenes de resonancia magnética funcional, fMRI) y biología molecular (mutaciones dirigidas al sitio) para estudiar la función cerebral.
A continuación aprenderá todo sobre
- Canales iónicos
- Membrana en reposo y potencial de acción
- Unión neuromuscular / sinapsis
- Conducción nerviosa
- Neurotransmisores, Receptores y vías
Canales iónicos
Un canal iónico es una macromolécula proteica que atraviesa la amplitud de una membrana y permite el paso de moléculas. Los iones se mueven en una dirección determinada por el gradiente electroquímico a través de la membrana.
- Los iones tienden a fluir desde una zona de alta concentración a una zona de baja concentración.
- En presencia de un gradiente de voltaje, puede no haber flujo de iones a pesar de las concentraciones desiguales.
- Los canales iónicos pueden estar abiertos o cerrados.
- La apertura se produce al cambiar el voltaje a través de la membrana, o al unir una sustancia química a un receptor.
- La función más importante es que proporcionan a la neurona excitabilidad eléctrica.
- Se encuentran en todas las partes de la neurona y en menor medida en las células neurogliales.
Tipos de canales
- Activados por voltaje
- Activados químicamente
- Estiramiento mecánico/presión.
Propiedades fundamentales de un canal iónico
- Está formado por una serie de subunidades proteicas, asentadas a lo largo de la membrana, que permiten a los iones cruzar de un lado a otro. (Poro transmembrana)
- El canal debe ser capaz de pasar del estado abierto al cerrado, y viceversa.
- Debe ser capaz de abrirse en respuesta a los estímulos adecuados.
Algunos canales responden a estímulos químicos (especialmente en la sinapsis). Estos canales tienen receptores específicos para esa sustancia química, que lleva a la apertura del canal.
Membrana en reposo y potencial de acción
- En el estado de reposo, la membrana de la célula neuronal es bastante impermeable a los iones. Esto es crucial para la generación del potencial de membrana en reposo.
- El principal ion intracelular es el potasio (es el sodio en el líquido extracelular).
- El flujo natural de iones por medio de sus gradientes de concentración es que el K+ salga de la célula y el Na+ entre.
- Este movimiento de iones fuera de la célula conduce a un potencial de membrana negativo – Hiperpolarización.
- Lo contrario ocurre con una entrada relativa de iones – Hipopolarización.

La membrana en reposo es relativamente impermeable a los iones Na+, mientras que permanece permeable a los iones K+. Así que cuando la membrana está en reposo, tenderá a haber un flujo de salida de iones K+ fuera de la célula, por su gradiente de concentración, dejando un exceso de carga negativa. Esto continúa hasta que el gradiente de concentración química que expulsa el K+ de la célula se encuentra exactamente en el punto en el que se compensa con la diferencia de potencial eléctrico generada por este efluvio (el potencial de membrana). (el potencial de membrana) El K+ vuelve a entrar en la célula.
Este estado estable de la membrana se denomina Potencial de Equilibrio.
La ecuación para derivar el potencial de equilibrio se llama ecuación de Nernst.
En realidad, en los axones hay una carga positiva ligeramente aumentada de forma inesperada debido a una pequeña permeabilidad a los iones Na+ de la membrana en el estado de reposo. Esto se compensa con la bomba de intercambio Na+/K+ dependiente del ATP. Bombea 3 iones de Na+ por cada 3 iones de K+ introducidos en la célula. Sin embargo, sólo hace una pequeña contribución al potencial de membrana en reposo de la célula.
Potencial de acción
Se define como un «impulso eléctrico único que pasa por un axón».
Es todo o nada en su acción. Esto significa que una vez alcanzada la intensidad umbral del estímulo, se generará un potencial de acción.
La información en el sistema nervioso se codifica e interpreta por la frecuencia de disparo, no por el tamaño del potencial de acción.
Umbral de intensidad del estímulo: El valor en el que la corriente neta de entrada (determinada por los iones de Na+) es justo mayor que la corriente neta de salida (llevada por los iones de K+). Normalmente es de -55mV (umbral crítico de disparo)
El PA se produce más fácilmente en el montículo del axón porque es donde hay una mayor densidad de canales de iones Na+. Por este motivo, éste es el lugar de inicio del PA en la neurona.
Si no se alcanza el umbral, no se genera el potencial de acción y la transmisión de la señal se termina en ese punto.
Secuencia de acontecimientos para que se genere un potencial de acción
- El voltaje de despolarización activa los canales iónicos de Na+ sensibles al voltaje en la membrana neuronal: Los iones Na+ fluyen por el gradiente electroquímico. La membrana se despolariza aún más. Se abren más canales de Na+ en un bucle de retroalimentación positiva. Cuando hay una mayor corriente de entrada de iones de sodio en comparación con la salida de iones de K+, se produce una rápida apertura de todos los canales de Na+. Esto despolariza la membrana hacia el potencial de equilibrio para el Na+ (+55mV). Se genera un pico de PA, pero no se alcanza el potencial de equilibrio para el Na+ debido al aumento del flujo de K+.
- A medida que los canales de Na+ se vuelven menos activos, el PA cae. Esta inactivación es dependiente del voltaje. Durante la fase de caída, la corriente de K+ es importante ya que conduce a un breve período de hiperpolarización de la membrana antes de que se desactive.
- El potencial de membrana vuelve al estado de reposo.
Unión Neuromuscular / Sinapsis
Sinapsis
Una sinapsis es la unión de dos neuronas. La sinapsis química es la que predomina en el sistema nervioso, sin embargo las sinapsis eléctricas se encuentran en el músculo cardíaco y en las células gliales.
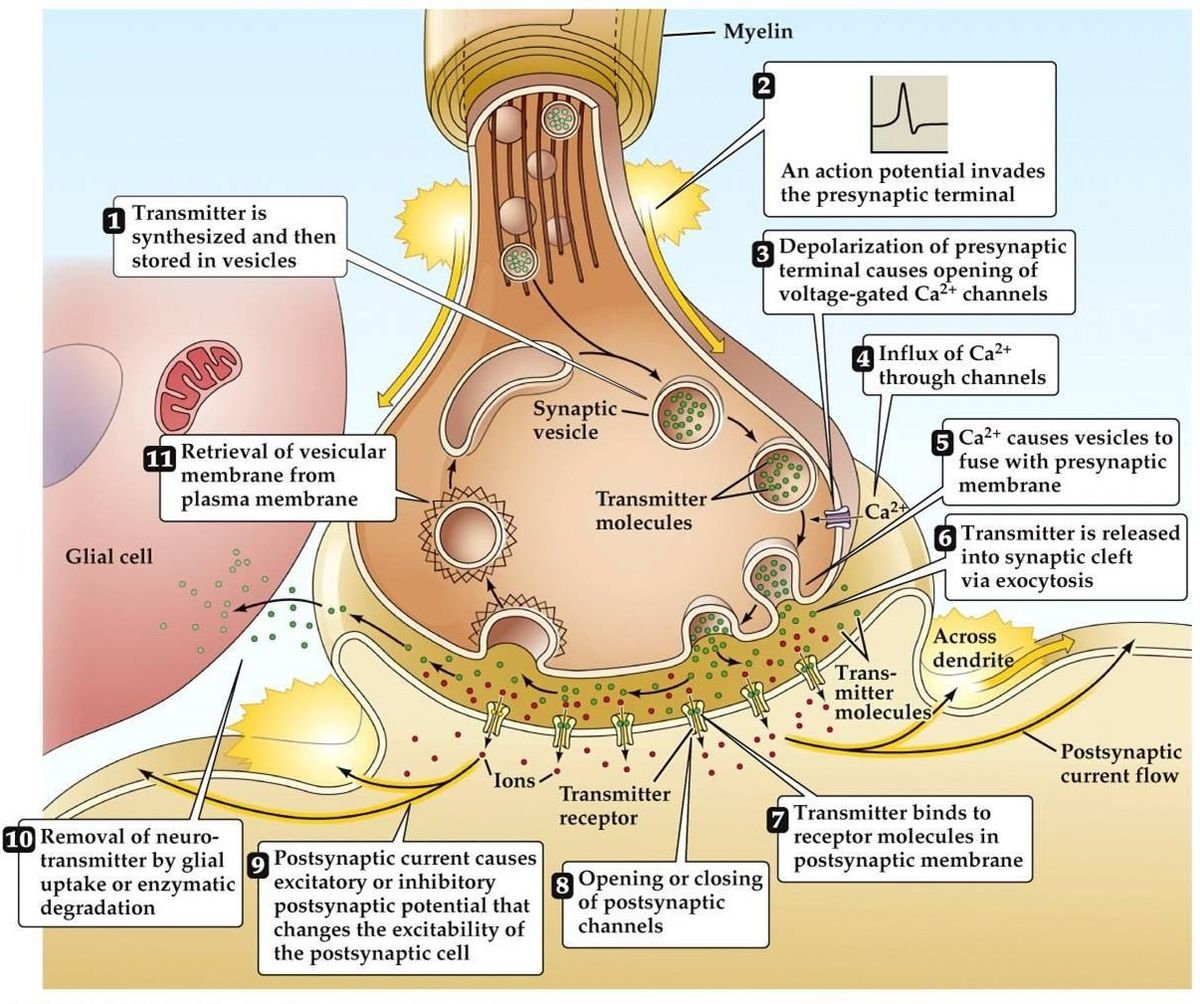
Transmisión sináptica
- Llega el potencial de acción, conduce a la despolarización del terminal presináptico. Los canales de Ca2+ dependientes del voltaje se abren en las zonas activas de la terminal. Esto conduce a la afluencia de Ca2+.
- El flujo de Ca2+ conduce a la fosforilación y a la alteración de la cantidad de proteínas de unión al calcio presinápticas. Esto libera la vesícula de su red de actina presináptica, Entonces se une a la membrana presináptica.
- La fusión de la vesícula con la membrana conduce a la formación de un pequeño canal, que se expande rápidamente y libera su contenido en la hendidura sináptica. La membrana de la vesícula se recicla por endocitosis.
- El neurotransmisor liberado se difunde a través de la hendidura sináptica con el objetivo de unirse al receptor postsináptico.
- La activación del receptor postsináptico conduce a un cambio en el potencial de la membrana postsináptica.
- Hay teorías alternativas que sugieren que el neurotransmisor se entrega a través de moléculas o canales de membrana en lugar de vesículas.
Conducción nerviosa
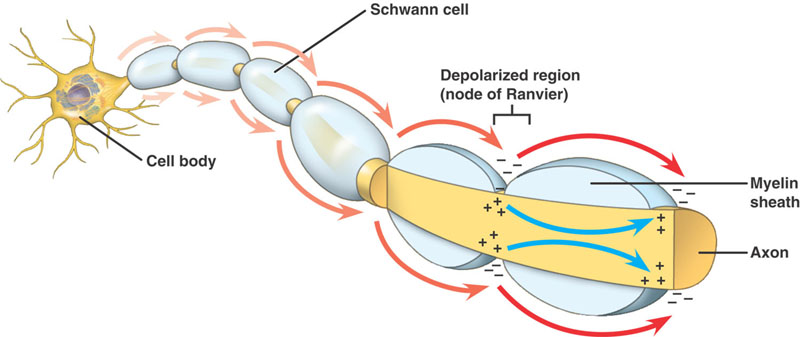
La propagación del potencial de acción se logra mediante la propagación local de la corriente.
El nervio está aislado con mielina si su tamaño es superior a un determinado diámetro, con los Nodos de Ranvier en varios intervalos a lo largo de su longitud.
Axones no mielinizados
El potencial de acción provoca la despolarización de la membrana inmediatamente anterior (y posterior). La membrana se encuentra en un estado refractario, por lo que el potencial de acción sólo se conduce en 1 dirección. Esto es factible en axones pequeños, pero la propagación de la corriente es lenta.
Axones mielinizados
La misma secuencia de eventos que en los axones no mielinizados. Sin embargo, hay una diferencia significativa. El potencial de acción que progresa se encuentra con una estructura de alta resistencia y baja capacidad: la mielina que envuelve al axón. La corriente despolarizante pasa a lo largo del axoplasma, hasta que alcanza el Nodo de Ranvier de baja resistencia con su gran cantidad de canales de Na+. El potencial de acción se genera entonces en este lugar. El potencial de acción se conduce de nodo a nodo. Esto se denomina conducción salatoria.
Ventajas de la mielinización
- Permite una rápida conducción del potencial de acción
- Minimiza las demandas metabólicas de la célula.
- Aumenta la capacidad de empaquetamiento del SN, lo que permite que haya más fibras en un nervio.
La mayoría de los nervios de más de 1 micra están mielinizados.
Las alteraciones de la conducción se deben normalmente a patologías de desmielinización. Entre ellas se encuentran el síndrome de Guillan Barre y la esclerosis múltiple.
Integración postsináptica:
Cada neurona central recibe muchos cientos de sinapsis. Cada una de estas entradas es luego integrada en una respuesta por esa neurona en particular. Esto implica el procesamiento de todas las entradas que llegan en un momento dado. Esto se denomina suma espacial. El procesamiento de las entradas durante un periodo de tiempo determinado se denomina suma temporal. Habrá una mezcla de sinapsis excitatorias e inhibitorias actuando en este punto.
Potenciales postsinápticos excitatorios
- La unión del neurotransmisor provoca la apertura de los canales iónicos.
- Hay afluencia de cationes en el proceso postsináptico
- Se produce la despolarización de la membrana.
- Las despolarizaciones registradas en la célula postsináptica ante un determinado estímulo excitatorio.
- Las despolarizaciones asociadas a los EPSPs pueden pasar a desencadenar APs si se produce la sumatoria en la sinapsis.
Potenciales Postsinápticos Inhibitorios
- Los canales iónicos permiten la entrada de aniones postsinápticos cuando están abiertos.
- Las hiperpolarizaciones de la membrana se producen debido a la entrada de CL- y a la salida de K+
- importante para modular la respuesta de las neuronas a la entrada excitatoria en la sinapsis.
- Se encuentran en sitios estratégicamente importantes de la neurona como la dendrita proximal y el soma.
- Tienen una gran influencia en el árbol dendrítico
- Algunas neuronas proporcionan su propia influencia inhibitoria al tener colaterales axónicos y sus propias interneuronas inhibidoras. Esto se denomina inhibición por retroalimentación. Por ejemplo, las motoneuronas y las células de Renshaw en la médula espinal.
Neurotransmisores, receptores y vías
El neurotransmisor se libera en la sinapsis y trabaja con una proteína específica en la membrana postsináptica llamada receptor. En algunas sinapsis, el neurotransmisor también puede interactuar con un autorreceptor presináptico. El PSA actúa como regulador de la cantidad de transmisor liberado.
Los receptores son normalmente específicos para un determinado neurotransmisor. Existen varios tipos de receptores. Los neurotransmisores liberados conjuntamente pueden regular la unión de otro transmisor.
Los receptores para neurotransmisores específicos pueden estar acoplados directamente a canales iónicos o a una enzima de membrana donde la unión del neurotransmisor al receptor abre un canal iónico a través de una cascada enzimática intracelular o modula indirectamente la probabilidad de que otros canales iónicos se abran en respuesta a cambios de voltaje. (neuromodulación)
El receptor activado sólo puede volver a su estado de reposo una vez que el neurotransmisor es eliminado por el proceso de hidrólisis enzimática (captación) en la terminal nerviosa presináptica o en las células gliales cercanas.
Desensibilización/baja regulación: Disminución de la afinidad del receptor por el transmisor a corto plazo, a largo plazo, esto conduce a una disminución del número de receptores.
Supersensibilidad/Regulación al alza: Un aumento de la afinidad del receptor por el transmisor a corto plazo puede conducir a un aumento de los canales a largo plazo.
Receptores:
- Ionotrópicos: N-metil-D-aspartato(NMDA)
- Sin NMDA.
- Metaotrópicos : Receptores de glutamato asociados a proteínas G que responden iniciando eventos bioquímicos intracelulares, modulando la transmisión sináptica.
Hay un gran número de Neurotransmisores:
- Aminoácidos excitatorios: Son los principales NT excitatorios del SNC. El principal NT de este grupo es el Glutamato que actúa en una serie de receptores.
- Aminoácidos inhibidores: Son los principales neurotransmisores inhibidores, el principal de los cuales es el GABA, presente en todo el SNC. La glicina, que se encuentra principalmente en la médula espinal.
- Monoaminas: Encontradas en pequeños grupos de neuronas en el tronco cerebral, proyectándose ampliamente en el SNC. Se encuentran en el SN autónomo. Se unen a una gran cantidad de receptores.
- Acetilcolina: Ampliamente distribuida por todo el sistema nervioso, incluyendo la unión neuromuscular y el SN autónomo.
- Neuropéptidos: Se encuentran en todo el SN. A menudo se liberan simultáneamente con otros NT.
Además, se han encontrado más de 50 péptidos neuroactivos, y regularmente se descubren otros nuevos. Muchos de ellos se «liberan conjuntamente» con un transmisor de molécula pequeña. Sin embargo, en algunos casos un péptido es el transmisor principal en una sinapsis. La β-endorfina es un ejemplo relativamente bien conocido de un neurotransmisor peptídico porque participa en interacciones altamente específicas con los receptores opioides en el sistema nervioso central.
Los iones simples (como el zinc liberado sinápticamente) también son considerados neurotransmisores por algunos, así como algunas moléculas gaseosas como el óxido nítrico (NO), el monóxido de carbono (CO) y el sulfuro de hidrógeno (H2S). Los gases se producen en el citoplasma neuronal y se difunden inmediatamente a través de la membrana celular al líquido extracelular y a las células cercanas para estimular la producción de segundos mensajeros. Los neurotransmisores gaseosos solubles son difíciles de estudiar, ya que actúan rápidamente y se descomponen de inmediato, existiendo sólo unos segundos.
El transmisor más frecuente es el glutamato, que es excitatorio en más del 90% de las sinapsis del cerebro humano. El siguiente más frecuente es el ácido gamma-aminobutírico, o GABA, que es inhibidor en más del 90% de las sinapsis que no utilizan glutamato. Aunque otros transmisores se utilizan en menos sinapsis, pueden ser muy importantes funcionalmente: la gran mayoría de las drogas psicoactivas ejercen sus efectos alterando las acciones de algunos sistemas de neurotransmisores, a menudo actuando a través de transmisores distintos del glutamato o el GABA. Las drogas adictivas como la cocaína y las anfetaminas ejercen sus efectos principalmente sobre el sistema dopaminérgico. Las drogas opiáceas adictivas ejercen sus efectos principalmente como análogos funcionales de los péptidos opioides, que, a su vez, regulan los niveles de dopamina.