Original Editor – Joanne Garvey and Naomi O’Reilly
Top Contributors – Joanne Garvey, Lucinda hampton, Rachael Lowe, Naomi O’Reilly and Tarina van der Stockt
- Introduction
- Introduction
- Esittely
- Ionikanavat
- Kanavatyypit
- Ionikanavan perusominaisuudet
- Levossa oleva kalvo ja toimintapotentiaali
- Aaktiopotentiaali
- Neuromuskulaarinen liitos / synapsit
- Synapsi
- Synaptinen transmissio
- Hermon johtuminen
- Myeliinittömät aksonit
- Myelinoidut aksonit
- Neurotransmitterit, reseptorit ja reitit
Introduction
Ensimmäinen toimittaja – Joanne Garvey ja Naomi O’Reilly
Introduction
Ensimmäinen toimittaja – Joanne Garvey, Lucinda hampton, Rachael Lowe, Naomi O’Reilly ja Tarina van der Stockt
Esittely
Neurofysiologian haara, joka käsittelee hermoelimistön tehtäviä. Eli hermosolujen, solukalvojen ja verkostojen toiminnallisten ominaisuuksien tutkiminen.
- Historiallisesti sitä on hallinnut elektrofysiologia – neuronaalisten tapahtumien sähköinen rekisteröinti aina molaarisesta (elektroenkefalogrammi, EEG) soluihin (yksittäisten neuronien ominaisuuksien solunsisäinen rekisteröinti).
- Koska neuroni on sähkökemiallinen kone, on mahdotonta erottaa sähköisiä tapahtumia biokemiallisista ja molekulaarisista prosesseista, jotka saavat ne aikaan.
- Neurofysiologit käyttävät nykyään kemian (kalsiumkuvantaminen), fysiikan (funktionaalinen magneettikuvaus, fMRI) ja molekyylibiologian (kohdekohtaiset mutaatiot) tekniikoita aivojen toiminnan tutkimiseen.
Alhaalla opit kaiken
- Ionikanavista
- Lepomembraanista ja toimintapotentiaalista
- Neuromuskulaarisesta liitoksesta / synapseista
- Hermojohtumisesta
- Neurotransmittereista, Reseptorit ja reitit
Ionikanava on proteiinimakromolekyyli, joka ylittää kalvon leveyden ja päästää molekyylit läpi. Ionit liikkuvat kalvon poikki kulkevan sähkökemiallisen gradientin määräämään suuntaan.
- Ionit pyrkivät virtaamaan korkean konsentraation alueelta matalan konsentraation alueelle.
- Jännitegradientin vallitessa ionien virtausta ei välttämättä tapahdu, vaikka pitoisuudet ovatkin epätasaisia.
- Ionikanavat voivat olla avoimia tai suljettuja.
- Avautuminen saadaan aikaan muuttamalla jännitettä kalvon yli tai sitomalla kemiallinen aine reseptoriin.
- Tärkein tehtävä on, että ne antavat hermosolulle sähköisen herätteen.
- Löytyy hermosolun kaikista osista ja vähäisemmässä määrin neurogliasoluista.
- Jänniteohjattu
- Kemiallisesti aktivoitu
- Mekaaninen venytys/paine.
- Kanava koostuu useista proteiinien alayksiköistä, jotka istuvat kalvon poikki ja sallivat ionien siirtymisen puolelta toiselle. (Transmembraanihuokoset)
- Kanavan on pystyttävä siirtymään avoimesta tilasta suljettuun tilaan ja takaisin.
- Pitää pystyä avautumaan vasteena sopiviin ärsykkeisiin.
Jotkut kanavat reagoivat kemiallisiin ärsykkeisiin (erityisesti synapsissa). Näillä kanavilla on spesifiset reseptorit kyseiselle kemikaalille, mikä johtaa kanavan avautumiseen.
Levossa oleva kalvo ja toimintapotentiaali
- Levossa olevassa tilassa hermosolun solukalvo on melko läpäisemätön ioneille. Tämä on ratkaisevaa lepomembraanipotentiaalin syntymisen kannalta.
- Solunsisäinen pääioni on kalium (solunulkoisessa nesteessä se on natrium).
- Ionien luonnollinen virtaus niiden konsentraatiogradienttien avulla on, että K+ poistuu solusta ja Na+ tulee soluun.
- Tämä ionien liike ulos solusta johtaa negatiiviseen kalvopotentiaaliin – hyperpolarisaatioon.
- Ionien suhteelliseen sisäänvirtaukseen pätee päinvastoin – Hypopolarisaatio.

Laakakalvo on suhteellisen läpäisemätön Na+ -ioneille, kun taas se pysyy läpäisevänä K+ -ioneille. Kun kalvo siis on levossa, K+-ioneja pyrkii virtaamaan ulos solusta sen konsentraatiogradienttia pitkin, jolloin ylimääräinen negatiivinen varaus jää jäljelle. Tämä jatkuu, kunnes kemiallinen konsentraatiogradientti, joka ajaa K+ -ionit ulos solusta, on täsmälleen siinä pisteessä, jossa tämän ulosvirtauksen aiheuttama sähköinen potentiaaliero kompensoi sen. (kalvopotentiaali) K+ vedetään sitten takaisin soluun.
Tätä kalvon tasaista tilaa kutsutaan tasapainopotentiaaliksi.
Tasapainopotentiaalin johtamisen yhtälöä kutsutaan Nernstin yhtälöksi.
Aksoneissa on itse asiassa hieman yllättäen lisääntynyt positiivinen varaus, joka johtuu kalvon pienestä läpäisevyydestä Na+ -ioneille lepotilassa. Tämän kompensoi ATP-riippuvainen Na+/K+-vaihtopumppu. Pumppaan ulos 3 Na+ -ionia jokaista soluun tuotua 3K+ -ionia kohden. Se vaikuttaa kuitenkin vain vähän solun lepokalvopotentiaaliin.
Aaktiopotentiaali
Määritellään ”yksittäiseksi sähköimpulssiksi, joka kulkee aksonia pitkin”.
Se on toiminnaltaan kaikki tai ei mitään. Tämä tarkoittaa, että kun ärsykkeen kynnysvoimakkuus saavutetaan, syntyy toimintapotentiaali.
Hermostossa informaatio koodataan ja tulkitaan laukaisutaajuudella, ei toimintapotentiaalin koolla.
Kynnysärsykkeen intensiteetti: Arvo, jolla sisäänpäin suuntautuva nettovirta (Na+ -ionien määräämä) on juuri suurempi kuin ulospäin suuntautuva nettovirta (K+ -ionien kuljettama). Se on normaalisti -55mV (kriittinen laukaisukynnys)
AP tapahtuu helpoimmin aksonikukkulassa, koska siellä Na+-ionikanavien tiheys on suurempi. Tästä syystä tämä on AP:n alkamispaikka neuronissa.
Jos kynnysarvoa ei saavuteta, aktiopotentiaalia ei synny, ja signaalin siirto loppuu siihen kohtaan.
Toimintapotentiaalin syntymisen tapahtumaketju
- Depolarisoiva jännite aktivoi hermosolukalvon jänniteherkät Na+ -ionikanavat: Na+ -ionit virtaavat sähkökemiallista gradienttia alaspäin. Kalvo depolarisoituu edelleen. Lisää Na+-kanavia aukeaa positiivisessa takaisinkytkennässä. Kun natriumionien sisäänvirtaus on suurempi kuin K+ -ionien ulosvirtaus, kaikki Na+ -kanavat avautuvat nopeasti. Tämä depolarisoi kalvon kohti Na+:n tasapainopotentiaalia (+55mV). Syntyy AP-piikki, mutta se ei saavuta Na+:n tasapainopotentiaalia K+:n lisääntyvän ulosvirtauksen vuoksi.
- Kun Na+ -kanavien aktiivisuus vähenee, AP laskee. Tämä inaktivoituminen on jännitteestä riippuvainen. Laskevan vaiheen aikana K+ -virta on tärkeä, koska se johtaa lyhyeen kalvon hyperpolarisaatiojaksoon ennen kuin se deaktivoituu.
- Membraanipotentiaali palaa lepotilaan.
Neuromuskulaarinen liitos / synapsit
Synapsi
Synapsi on kahden hermosolun liitoskohta. Kemiallinen synapsi on vallitseva hermostossa esiintyvä synapsi, mutta sähköisiä synapseja esiintyy sydänlihaksessa ja gliasoluissa.
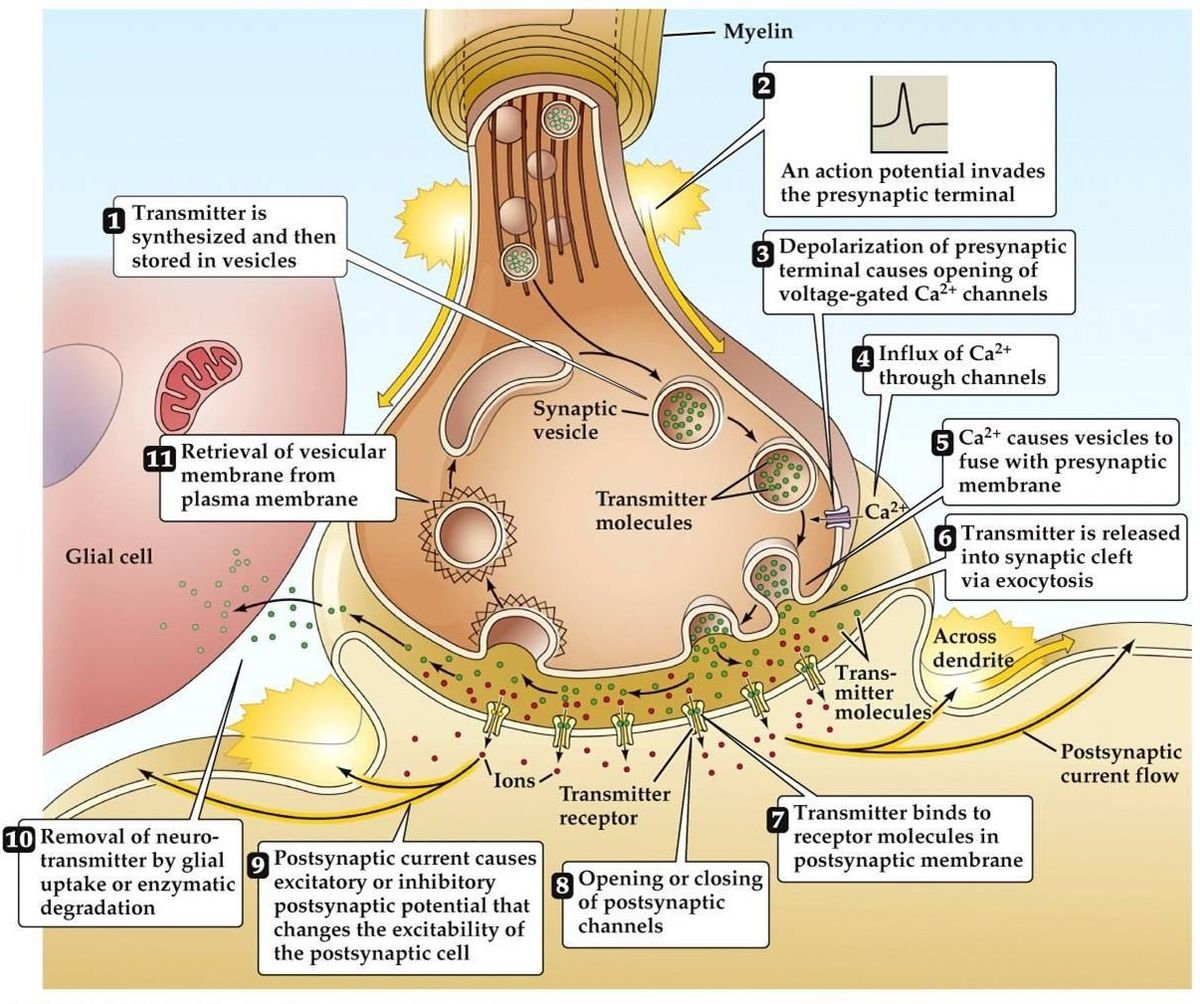
Synaptinen transmissio
- Aktiopotentiaali saapuu, johtaa presynaptisen päätteen depolarisaatioon. Jännitteestä riippuvat Ca2+-kanavat avautuvat terminaalin aktiivisilla vyöhykkeillä. Tämä johtaa Ca2+:n sisäänvirtaukseen.
- Ca2+:n sisäänvirtaus johtaa presynaptisten kalsiumia sitovien proteiinien fosforylaatioon ja määrän muuttumiseen. Tämä vapauttaa vesikkelin presynaptisesta aktiiniverkostosta, Se sitoutuu sitten presynaptiseen kalvoon.
- Vesikkelin sulautuminen kalvoon johtaa pienen kanavan muodostumiseen, joka laajenee nopeasti ja vapauttaa sisältönsä synaptiseen rakoon. Vesikkelin kalvo kierrätetään endosytoosin avulla.
- Vapautunut välittäjäaine diffundoituu synaptisen raon läpi tavoitteenaan sitoutua postsynaptiseen reseptoriin.
- Postsynaptisen reseptorin aktivoituminen johtaa postsynaptisen kalvopotentiaalin muutokseen.
- On olemassa vaihtoehtoisia teorioita, joiden mukaan välittäjäaine kulkeutuu pikemminkin molekyylien tai kalvokanavien kuin vesikkelien kautta.
Hermon johtuminen
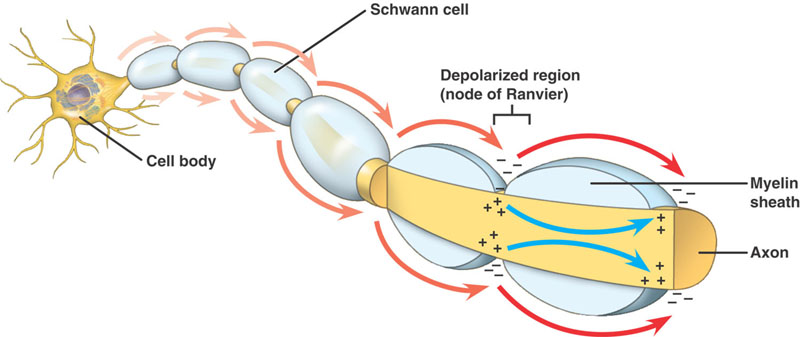
Toimintapotentiaalin eteneminen tapahtuu paikallisen virran leviämisen avulla.
Hermo on eristetty myeliinillä, jos sen koko ylittää tietyn halkaisijan, ja sen pituudella on eri välein Ranvierin solmukohtia.
Myeliinittömät aksonit
Toimintapotentiaali johtaa välittömästi edessä (ja takana) olevan kalvon depolarisaatioon. Kalvo on refraktorisessa tilassa, joten toimintapotentiaali johdetaan vain yhteen suuntaan. Tämä on toteutettavissa pienissä aksoneissa, mutta virran leviäminen on hidasta.
Myelinoidut aksonit
Sama tapahtumasarja kuin myelinoimattomissa aksoneissa. Siinä on kuitenkin merkittävä ero. Etenevä aktiopotentiaali kohtaa korkean resistanssin, matalan kapasitanssin rakenteen – aksonin ympärille kietoutuneen myeliinin. Depolarisoituva virta kulkee aksoplasmaa pitkin, kunnes se saavuttaa matalaresistanssisen Ranvierin solmun, jossa on suuri määrä Na+ -kanavia. Toimintapotentiaali syntyy sitten tässä kohdassa. Toimintapotentiaali johdetaan solmusta solmuun. Tätä kutsutaan Saltatory Conduction -johtumiseksi.
Myelinaation edut
- Mahdollistaa toimintapotentiaalin nopean johtumisen
- Minimoi solun metaboliset vaatimukset.
- Lisää NS:n pakkautumiskykyä, jolloin yhteen hermoon mahtuu enemmän kuituja.
Useimmat yli 1 mikronin hermot ovat myelinoidut.
Johtumishäiriöt johtuvat yleensä demyelinaatiopatologioista. Niitä ovat esimerkiksi Guillan Barren oireyhtymä ja multippeliskleroosi.
Postsynaptinen integraatio:
Kukin keskushermosolu saa useita satoja synapseja. Kukin näistä syötteistä integroidaan sitten kyseisen neuronin vasteeksi. Tämä edellyttää kaikkien kerrallaan saapuvien syötteiden käsittelyä. Tätä kutsutaan spatiaaliseksi summaukseksi. Tietyn ajanjakson aikana tapahtuvaa syötteiden käsittelyä kutsutaan ajalliseksi summautumiseksi. Tässä vaiheessa toimii sekä kiihottavia että estäviä synapseja.
Excitatoriset postsynaptiset potentiaalit
- Neurotransmitterin sitoutuminen johtaa ionikanavien avautumiseen.
- Post-synaptisessa prosessissa tapahtuu kationien sisäänvirtaus
- Tapahtuu kalvon depolarisaatio.
- EPSP:t depolarisaatioita, jotka rekisteröidään postsynaptisessa solussa tiettyyn eksitatoriseen ärsykkeeseen.
- EPSP:iin assosioituvat depolarisaatiot voivat jatkossa laukaista AP:t, jos synapsissa tapahtuu summautumista.
Inhibitoriset postsynaptiset potentiaalit
- Ionikanavat mahdollistavat postsynaptisen anionin sisäänvirtauksen ollessaan auki.
- Kalvon hyperpolarisoituminen tapahtuu CL-:n sisäänvirtauksen ja K+:n ulosvirtauksen vuoksi
- tärkeää hermosolujen vasteen moduloinnissa synapsissa tapahtuvaan eksitatoriseen syötteeseen.
- Löytyy neuronin strategisesti tärkeissä paikoissa, kuten proksimaalisessa dendriitissä ja somassa.
- Neuroneilla on suuri vaikutus dendriittipuuhun
- Jotkut neuronit tarjoavat oman inhiboivan vaikutuksensa sillä, että niillä on aksonikollateraaleja ja omia inhiboivia interneuroneja. Tätä kutsutaan takaisinkytkentäinhibitioksi. Esim. motorneuronit ja Renshaw-solut selkäytimessä.
Neurotransmitterit, reseptorit ja reitit
Neurotransmitteri vapautuu synapsissa ja toimii postsynaptisessa kalvossa olevan tietyn proteiinin, reseptorin, kanssa. Joissakin synapseissa välittäjäaine voi olla vuorovaikutuksessa myös presynaptisen autoreseptorin kanssa. PSA toimii vapautuvan lähettimen määrän säätelijänä.
Reseptorit ovat yleensä spesifisiä tietylle välittäjäaineelle. Reseptoreita on useita erilaisia. Yhdessä vapautuvat välittäjäaineet voivat säädellä toisen välittäjäaineen sitoutumista.
Spesifisten välittäjäaineiden reseptorit voivat olla joko kytketty suoraan ionikanaviin tai kalvoentsyymiin, jolloin välittäjäaineen sitoutuminen reseptoriin joko avaa ionikanavan solunsisäisen entsyymikaskadin kautta tai moduloi epäsuorasti muiden ionikanavien avautumistodennäköisyyttä vasteena jännitteen muutoksiin. (neuromodulaatio)
Aktivoitunut reseptori voi palata lepotilaansa vasta, kun välittäjäaine on poistunut entsymaattisen hydrolyysiprosessin (uptake) kautta presynaptiseen hermopäätteeseen tai läheisiin gliasoluihin.
Desensitisaatio/alasäätely: Reseptorin affiniteetin väheneminen lähetintä kohtaan lyhyellä aikavälillä, pitkällä aikavälillä tämä johtaa reseptorien määrän vähenemiseen.
Yliherkkyys/ylössäätely: Reseptorin lisääntynyt affiniteetti lähetintä kohtaan lyhyellä aikavälillä voi johtaa kanavien lisääntymiseen pitkällä aikavälillä.
Reseptorit:
- Ionotrooppiset: N-metyyli-D-aspartaatti (NMDA)
- Ei NMDA.
- Metabotrooppiset : G-proteiiniin assosioituneet glutamaattireseptorit, jotka reagoivat käynnistämällä solunsisäisiä biokemiallisia tapahtumia, jotka moduloivat synaptista transmissiota.
Neurotransmittereita on suuri määrä:
- Eksitatoriset aminohapot: Nämä ovat CNS:n tärkeimmät eksitatoriset NT:t. Tärkein NT tässä ryhmässä on glutamaatti, joka vaikuttaa useisiin reseptoreihin.
- Inhibitoriset aminohapot: Nämä ovat tärkeimmät inhibitoriset välittäjäaineet, joista tärkein on GABA, jota esiintyy koko keskushermostossa. Glysiini, jota esiintyy pääasiassa selkäytimessä.
- Monoamiinit: Löytyvät aivorungon pienistä neuroniryhmistä, jotka projisoituvat laajasti keskushermostoon. Löytyy autonomisesta NS:stä. Ne sitoutuvat lukuisiin reseptoreihin.
- Asetyylikoliini: Levinnyt laajalti koko hermostoon, mukaan lukien neuromuskulaarinen liitos ja autonominen NS.
- Neuropeptidit: Löytyy kaikkialla NS:ssä. Vapautuvat usein samanaikaisesti muiden NT:iden kanssa.
Lisäksi on löydetty yli 50 neuroaktiivista peptidiä, ja uusia löydetään säännöllisesti. Monet näistä ”vapautuvat” yhdessä pienimolekyylisen lähettimen kanssa. Joissakin tapauksissa peptidi on kuitenkin ensisijainen lähetin synapsissa. β-endorfiini on suhteellisen tunnettu esimerkki peptidivälittäjäaineesta, koska se on hyvin spesifisessä vuorovaikutuksessa keskushermoston opioidireseptorien kanssa.
Jotkut pitävät myös yksittäisiä ioneja (kuten synaptisesti vapautuvaa sinkkiä) neurotransmittereina, samoin kuin joitakin kaasumaisia molekyylejä, kuten typpioksidia (NO), hiilimonoksidia (CO) ja rikkivetyä (H2S). Kaasuja tuotetaan hermosolujen sytoplasmassa ja ne diffundoituvat välittömästi solukalvon läpi solunulkoiseen nesteeseen ja läheisiin soluihin stimuloimaan toisten sanansaattajien tuotantoa. Liukoisia kaasun välittäjäaineita on vaikea tutkia, koska ne vaikuttavat nopeasti ja hajoavat välittömästi, sillä ne ovat olemassa vain muutaman sekunnin ajan.
Yleisin lähetin on glutamaatti, joka kiihdyttää reilusti yli 90 prosenttia ihmisaivojen synapseista. Seuraavaksi yleisin on gamma-aminovoihappo eli GABA, joka on inhiboiva yli 90 prosentissa synapseista, jotka eivät käytä glutamaattia. Vaikka muita välittäjäaineita käytetään harvemmissa synapseissa, ne voivat olla toiminnallisesti hyvin tärkeitä: suurin osa psykoaktiivisista lääkkeistä vaikuttaa muuttamalla joidenkin välittäjäainejärjestelmien toimintaa, usein muiden välittäjäaineiden kuin glutamaatin tai GABA:n kautta. Kokaiinin ja amfetamiinin kaltaiset riippuvuutta aiheuttavat huumeet vaikuttavat pääasiassa dopamiinijärjestelmään. Riippuvuutta aiheuttavat opiaattilääkkeet vaikuttavat ensisijaisesti opioidipeptidien toiminnallisina analogeina, jotka puolestaan säätelevät dopamiinitasoja.