Objectifs d’apprentissage
- Dire les trois parties fondamentales d’un désoxyribonucléotide.
- Dire quelles bases azotées sont des purines et quelles sont des pyrimidines.
- Définir l’appariement complémentaire des bases.
- Dire pourquoi l’ADN ne peut être synthétisé que dans le sens 5′ vers 3′.
- Comparer le nucléoïde procaryote au noyau eucaryote en fonction des éléments suivants :
- nombre de chromosomes
- chromosomes linéaires ou circulaires
- présence ou absence d’une membrane nucléaire
- présence ou absence de nucléosomes
- présence ou absence de mitose
- présence ou absence de méiose
L’ADN est une longue, double brin, molécule hélicoïdale composée de blocs de construction appelés désoxyribonucléotides. Chaque désoxyribonucléotide est composé de trois parties : une molécule de désoxyribose, un sucre à 5 carbones, une base azotée et un groupe phosphate (figure \(\PageIndex{1}\)).
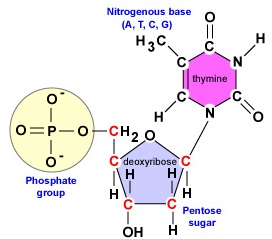
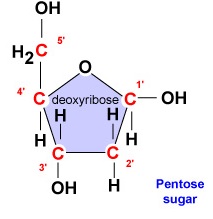
- Désoxyribose. Le désoxyribose est un sucre à 5 carbones annelés (figure \(\PageIndex{2}\)). Les 5 carbones sont numérotés séquentiellement dans le sens des aiguilles d’une montre autour du sucre. Les 4 premiers carbones forment en fait l’anneau du sucre, le carbone 5′ se détachant du carbone 4′ de l’anneau. La base azotée du nucléotide est fixée au carbone 1′ du sucre et le groupe phosphate est lié au carbone 5′. Lors de la synthèse de l’ADN, le groupe phosphate d’un nouveau désoxyribonucléotide est fixé de manière covalente par l’enzyme ADN polymérase au carbone 3′ d’un nucléotide déjà présent dans la chaîne.
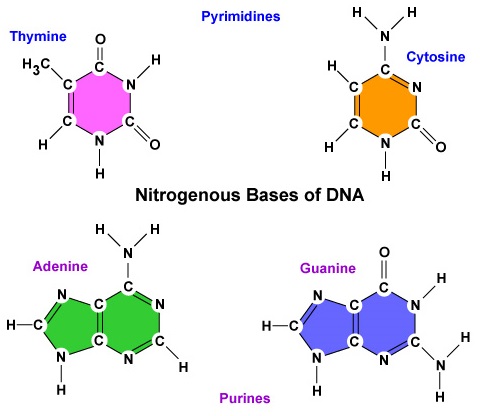
- Une base azotée. On trouve quatre bases azotées dans l’ADN : l’adénine, la guanine, la cytosine ou la thymine. L’adénine et la guanine sont connues comme des bases puriques tandis que la cytosine et la thymine sont connues comme des bases pyrimidiques (figure \(\PageIndex{3}\)).
- Un groupe phosphate (figure \(\PageIndex{4}\)).
Pour synthétiser les deux chaînes de désoxyribonucléotides lors de la réplication de l’ADN, les enzymes ADN polymérases impliquées sont seulement capables de joindre le groupe phosphate du carbone 5′ d’un nouveau nucléotide au groupe hydroxyle (OH) du carbone 3′ d’un nucléotide (Figure \(\PageIndex{2}\)) déjà dans la chaîne. La liaison covalente qui unit les nucléotides s’appelle une liaison phosphodiester. Chaque brin d’ADN possède ce que l’on appelle une extrémité 5′ et une extrémité 3′. Cela signifie qu’une extrémité de chaque brin d’ADN, appelée extrémité 5′, aura toujours un groupe phosphate attaché au carbone 5′ de son désoxyribonucléotide terminal (Figure \(\PageIndex{5}\)). L’autre extrémité de ce brin, appelée extrémité 3′, aura toujours un hydroxyle (OH) sur le carbone 3′ de son désoxyribonucléotide terminal.
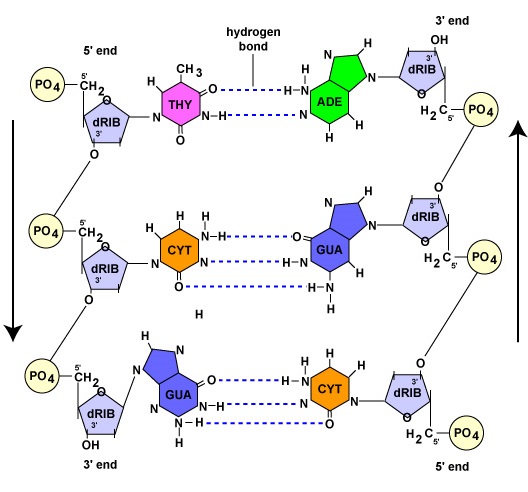
Comme nous le verrons dans la section suivante, chaque brin parent, lors de la réplication de l’ADN, sert de matrice pour la synthèse de l’autre brin par le biais de l’appariement complémentaire des bases. L’appariement complémentaire des bases fait référence aux nucléotides de l’ADN dont la base est l’adénine, qui ne forment des liaisons hydrogène qu’avec les nucléotides dont la base est la thymine (A-T). De même, les nucléotides dont la base est la guanine ne peuvent former des liaisons hydrogène qu’avec les nucléotides dont la base est la cytosine (G-C). (Dans le cas des nucléotides de l’ARN, comme nous le verrons plus loin, les nucléotides de l’adénine forment des liaisons hydrogène avec les nucléotides ayant la base uracile puisque la thymine n’est pas présente dans l’ARN). En raison de cette liaison, l’ADN prend sa forme hélicoïdale. Par conséquent, les deux brins d’ADN sont dits complémentaires. Partout où un brin possède un nucléotide contenant de l’adénine, le brin opposé aura toujours un nucléotide de thymine ; partout où il y a un nucléotide contenant de la guanine, le brin opposé aura toujours un nucléotide de cytosine (figure \(\PageIndex{1}\)).
Bien que les deux brins d’ADN soient complémentaires, ils sont orientés dans des directions opposées l’un à l’autre. On dit d’un brin qu’il est orienté de 5′ en 3′ ; le brin d’ADN opposé est antiparallèle, ou de 3′ en 5′ (figure \(\PageIndex{1}\)).
Nous allons maintenant comparer brièvement le génome des cellules procaryotes à celui des cellules eucaryotes.
Le génome procaryote (bactérien)
La zone au sein d’une bactérie où le chromosome peut être vu au microscope électronique est appelée nucléoïde. Le chromosome de la plupart des procaryotes est généralement une longue et unique molécule d’ADN double brin, hélicoïdale et super enroulée qui forme un cercle physique et génétique. Le chromosome mesure généralement environ 1000 µm de long et contient fréquemment près de 4000 gènes (Figure \(\PageIndex{8}\)). Escherichia coli, qui mesure 2 à 3 µm de long, possède un chromosome d’environ 1400 µm de long. Pour permettre à une macromolécule de cette taille de tenir dans la bactérie, des protéines de type histone se lient à l’ADN, séparant la molécule d’ADN en une cinquantaine de domaines chromosomiques et la rendant plus compacte. Une enzyme ADN topoisomérase appelée ADN gyrase super enroule ensuite le chromosome en un faisceau serré formant une masse d’ADN compacte et super enroulée d’environ 0,2 µm de diamètre.
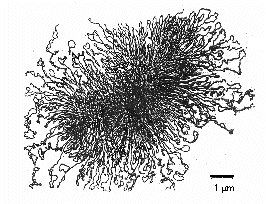
Les enzymes bactériennes appelées ADN topoisomérases sont essentielles au déroulement, à la réplication et au réenroulement de l’ADN bactérien circulaire et super enroulé (figure \(\PageIndex{7}\)). Ils sont également essentiels dans la transcription de l’ADN en ARN, dans la réparation de l’ADN et dans la recombinaison génétique chez les bactéries.
Figure \(\PageIndex{7}\) : ADN procaryote circulaire et superenroulé. Pour permettre à la grande molécule d’ADN de tenir dans la bactérie, une enzyme ADN topoisomérase appelée ADN gyrase super enroule le chromosome en un faisceau serré formant une masse d’ADN compacte et super enroulée d’environ 0,2 µm de diamètre.
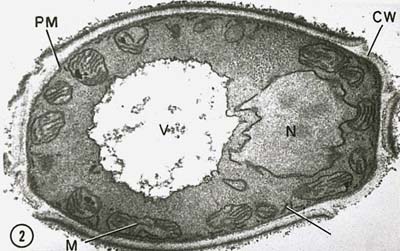
Le nucléoïde procaryote n’a pas de membrane nucléaire entourant l’ADN et le corps nucléaire ne se divise pas par mitose. La membrane cytoplasmique joue un rôle dans la séparation de l’ADN pendant la réplication bactérienne. Comme les bactéries sont haploïdes (n’ont qu’un seul chromosome), il n’y a pas non plus de méiose.
Le génome eucaryote
Les cellules procaryotes et eucaryotes diffèrent de façon très détaillée à la fois par la quantité et l’organisation de leurs molécules d’ADN. Les cellules eucaryotes contiennent beaucoup plus d’ADN que les bactéries, et cet ADN est organisé sous forme de multiples chromosomes situés à l’intérieur d’un noyau.
Le noyau des cellules eucaryotes est entouré d’une membrane nucléaire (figure \(\PageIndex{7}\)) et contient des chromosomes linéaires composés d’ADN chargé négativement associés à des protéines de base chargées positivement appelées histones pour former des structures appelées nucléosomes. Les nucléosomes font partie de ce que l’on appelle la chromatine, c’est-à-dire l’ADN et les protéines qui constituent les chromosomes. Le noyau se divise ma mitose et les cellules sexuelles haploïdes sont produites à partir de cellules diploïdes par la méiose.
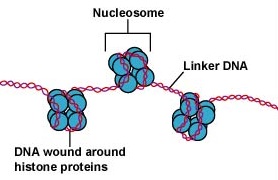
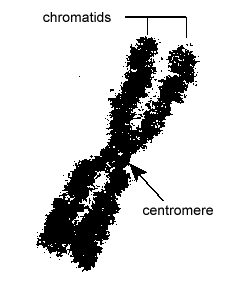
L’ADN des cellules eucaryotes est emballé de manière très organisée. Il est constitué d’une unité de base appelée nucléosome, une structure en forme de perle de 11 nm de diamètre qui se compose de 146 paires de bases d’ADN enroulées autour de huit molécules d’histones. Les nucléosomes sont reliés entre eux par un segment d’ADN d’environ 60 paires de bases appelé ADN lieur (Figure \(\PageIndex{9}\)). Une autre histone associée à l’ADN lieur emballe ensuite les nucléotides adjacents pour former un fil de nucléosomes de 30 nm de diamètre. Enfin, ces fils de nucléosomes emballés forment de grandes boucles enroulées qui sont maintenues ensemble par des protéines d’échafaudage non histones. Ces boucles enroulées sur les protéines d’échafaudage interagissent pour former la chromatine condensée observée dans les chromosomes pendant la mitose (Figure \(\PageIndex{10}\)).
Ces dernières années, on a découvert que la nature structurelle de la désoxyribonucléoprotéine contribue à la transcription ou non de l’ADN en ARN. Par exemple, des modifications chimiques de la chromatine peuvent permettre à des parties de celle-ci de se condenser ou de se détendre. Lorsqu’une région est condensée, les gènes ne peuvent pas être transcrits. En outre, des produits chimiques peuvent se fixer aux protéines histones autour desquelles l’ADN s’enroule ou en être retirés. La fixation ou le retrait de ces groupes chimiques sur l’histone détermine si l’expression des gènes à proximité est amplifiée ou réprimée.
L’épigénome fait référence à une variété de composés chimiques qui modifient le génome généralement en ajoutant un groupe méthyle (CH3) à la base nucléotidique adénine à des endroits spécifiques le long de la molécule d’ADN. Cette méthylation peut, à son tour, réprimer ou activer la transcription de gènes spécifiques. En activant ou désactivant les gènes, l’épigénome permet au génome d’interagir avec l’environnement de la cellule et d’y répondre. L’épigénome peut être hérité tout comme le génome.
Sommaire
- L’acide désoxyribonucléique (ADN) est une longue molécule hélicoïdale à double brin composée de blocs de construction appelés désoxyribonucléotides.
- Un désoxyribonucléotide est composé de 3 parties : une molécule de désoxyribose, un sucre à 5 carbones, une base azotée et un groupe phosphate.
- On trouve quatre bases azotées dans l’ADN : l’adénine, la guanine, la cytosine ou la thymine. L’adénine et la guanine sont connues comme des bases puriques tandis que la cytosine et la thymine sont connues comme des bases pyrimidiques.
- Le désoxyribose est un sucre annelé à 5 carbones. Les 5 carbones sont numérotés séquentiellement dans le sens des aiguilles d’une montre autour du sucre. Les 4 premiers carbones forment en fait l’anneau du sucre, le carbone 5′ se détachant du carbone 4′ de l’anneau. La base azotée du nucléotide est fixée au carbone 1′ du sucre et le groupe phosphate est lié au carbone 5′.
- Lors de la synthèse de l’ADN, l’enzyme ADN polymérase ne peut fixer le groupe phosphate d’un nouveau désoxyribonucléotide qu’au carbone 3′ d’un nucléotide déjà présent dans la chaîne.
- Lors de la réplication de l’ADN, chaque brin parent sert de matrice pour la synthèse de l’autre brin par le biais de l’appariement complémentaire des bases.
- L’appariement complémentaire des bases fait référence aux nucléotides de l’ADN ayant la base adénine ne formant des liaisons hydrogène qu’avec les nucléotides ayant la base thymine (A-T). De même, les nucléotides ayant la base guanine ne peuvent se lier par hydrogène qu’avec les nucléotides ayant la base cytosine (G-C).
- Alors que les deux brins d’ADN sont complémentaires, ils sont orientés dans des directions opposées l’un à l’autre. On dit d’un brin qu’il est orienté de 5′ à 3′ ; le brin d’ADN opposé est antiparallèle, ou de 3′ à 5′.
- Dans les cellules procaryotes, il n’y a pas de membrane nucléaire entourant l’ADN. Les cellules procaryotes sont dépourvues de mitose et de méiose.
- Pour permettre à une macromolécule aussi grande de tenir dans la bactérie, des protéines de type histone se lient à l’ADN, ségrégeant la molécule d’ADN en une cinquantaine de domaines chromosomiques et la rendant plus compacte. Ensuite, une enzyme appelée ADN gyrase enroule chaque domaine autour d’elle-même pour former une masse d’ADN compacte et super enroulée. Une topoisomérase appelée ADN gyrase catalyse le superenroulement négatif de l’ADN circulaire présent chez les bactéries. La topoisomérase IV, quant à elle, est impliquée dans la relaxation de l’ADN circulaire superenroulé, permettant la séparation des chromosomes filles reliés entre eux à la fin de la réplication de l’ADN bactérien.
- L’ADN des cellules eucaryotes est emballé dans des unités de base appelées nucléosomes, une structure en forme de perle constituée d’ADN enroulé autour de huit molécules d’histones. L’ADN est organisé en plusieurs chromosomes situés dans un noyau entouré d’une membrane nucléaire. Le noyau se divise par mitose et les gamètes sont produits par méiose chez les eucaryotes à reproduction sexuée.
- La nature structurelle de la désoxyribonucléoprotéine contribue à la transcription ou non de l’ADN en ARN. Par exemple, des modifications chimiques de la chromatine peuvent permettre à des portions de celle-ci de se condenser ou de se détendre. Lorsqu’une région est condensée, les gènes ne peuvent pas être transcrits. En outre, des produits chimiques peuvent s’attacher aux protéines histones autour desquelles l’ADN s’enroule ou en être retirés. La fixation ou le retrait de ces groupes chimiques à l’histone détermine si l’expression des gènes à proximité est amplifiée ou réprimée.
Contributeurs et attributions
-
Dr Gary Kaiser (COMMUNITY COLLEGE OF BALTIMORE COUNTY, CATONSVILLE CAMPUS)
.