Læringsmål
- Angiv de tre grundlæggende dele af et deoxyribonukleotid.
- Oplys, hvilke nitrogenbaser der er puriner, og hvilke der er pyrimidiner.
- Beskriv komplementær baseparring.
- Oplys, hvorfor DNA kun kan syntetiseres i 5′- til 3′-retningen.
- Sammenlign den prokaryote nukleoid med den eukaryote kerne med hensyn til følgende:
- antal kromosomer
- lineære eller cirkulære kromosomer
- forekomst eller fravær af en kernemembran
- forekomst eller fravær af nukleosomer
- forekomst eller fravær af mitose
- forekomst eller fravær af meiose
DNA er en lang, dobbeltstrenget, spiralformet molekyle, der består af byggesten kaldet desoxyribonukleotider. Hvert deoxyribonukleotid består af tre dele: et molekyle af det 5-carbonede sukkerstof deoxyribose, en nitrogenbas og en fosfatgruppe (figur \(\(\PageIndex{1}\)).
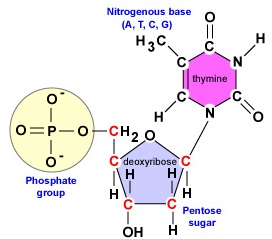
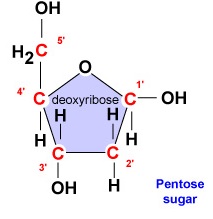
- Deoxyribose. Deoxyribose er et ringformet 5-kulstofsukker (figur \(\PageIndex{2}\)). De 5 kulstofatomer er nummereret i rækkefølge med uret rundt om sukkeret. De første 4 kulstofatomer danner faktisk sukkerets ring, idet 5′-kulstoffet kommer fra 4′-kulstoffet i ringen. Nukleotidets nitrogenbase er bundet til sukkerets 1′-kulstof, og fosfatgruppen er bundet til 5′-kulstoffet. Under DNA-syntesen bindes fosfatgruppen i et nyt desoxyribonukleotid kovalent af enzymet DNA-polymerase til 3′-kulstoffet i et nukleotid, der allerede findes i kæden.
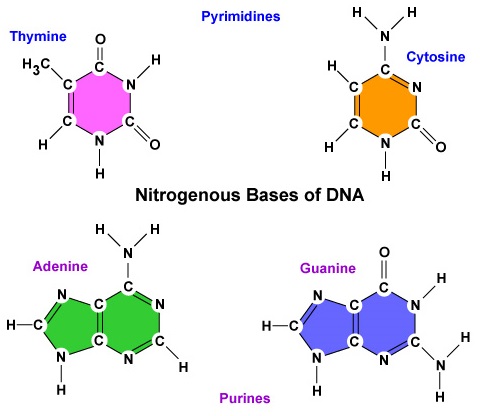
- En nitrogenholdig base. Der findes fire nitrogenbaserede baser i DNA: adenin, guanin, cytosin eller thymin. Adenin og guanin er kendt som purinbaser, mens cytosin og thymin er kendt som pyrimidinbaser (figur \(\(\PageIndex{3}\))).
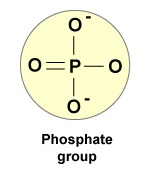
- En fosfatgruppe (figur \(\(\PageIndex{4}\)).
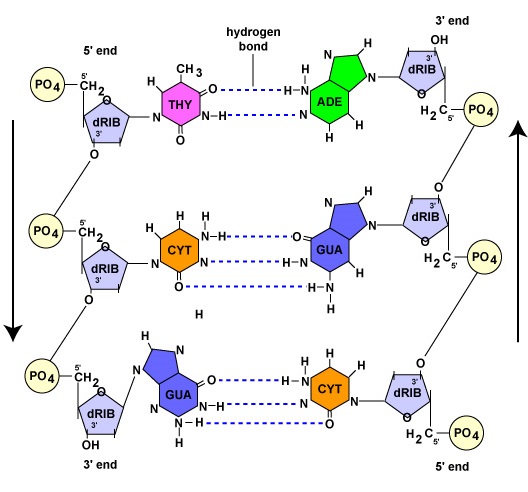
Som det vil fremgå af næste afsnit, fungerer hver moderstreng under DNA-replikationen som skabelon for syntesen af den anden streng ved hjælp af komplementær baseparring. Komplementær baseparring henviser til, at DNA-nukleotider med basen adenin kun danner hydrogenbindinger med nukleotider, der har basen thymin (A-T). På samme måde kan nukleotider med basen guanin kun hydrogenbindes med nukleotider med basen cytosin (G-C). (I tilfælde af RNA-nukleotider danner adeninnukleotider, som det vil blive vist senere, hydrogenbindinger med nukleotider med basen uracil, da thymin ikke findes i RNA). Som følge af denne binding antager DNA’et sin helikale form. Man siger derfor, at de to DNA-strenge er komplementære. Hvor den ene streng har et adeninholdigt nukleotid, vil den modsatte streng altid have et thymin-nukleotid; hvor der er et guaninholdigt nukleotid, vil den modsatte streng altid have et cytosin-nukleotid (figur \(\PageIndex{1}\)).
Selv om de to DNA-strenge er komplementære, er de orienteret i modsatte retninger i forhold til hinanden. Den ene streng siges at løbe 5′ til 3′; den modsatte DNA-streng løber antiparallelt, eller 3′ til 5′ (Figur \(\PageIndex{1}\)).
Vi vil nu kort sammenligne genomet i prokaryote celler med genomet i eukaryote celler.
Det prokaryote (bakterielle) genom
Det område i en bakterie, hvor kromosomet kan ses med et elektronmikroskop, kaldes en nukleoid. Kromosomet hos de fleste prokaryoter er typisk ét langt, enkelt molekyle af dobbeltstrenget, spiralformet, superviklet DNA, som danner en fysisk og genetisk cirkel. Kromosomet er generelt omkring 1000 µm langt og indeholder ofte omkring 4000 gener (Figur \(\PageIndex{8}\)). Escherichia coli, som er 2-3 µm lang, har et kromosom, der er ca. 1400 µm langt. For at et så stort makromolekyle kan få plads i bakterien, binder histonlignende proteiner sig til DNA’et og opdeler DNA-molekylet i ca. 50 kromosomale domæner, hvilket gør det mere kompakt. Et DNA-topoisomeraseenzym kaldet DNA-gyrase supervikler derefter kromosomet til et tæt bundt og danner en komprimeret, superviklet masse af DNA med en diameter på ca. 0,2 µm.
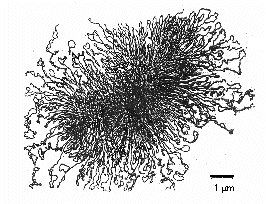
Bakterielle enzymer kaldet DNA-topoisomeraser er afgørende for afviklingen, replikationen og genopviklingen af det cirkulære, superoprullede bakterielle DNA (Figur \(\PageIndex{7}\)). De er også vigtige ved transkription af DNA til RNA, ved DNA-reparation og ved genetisk rekombination i bakterier.
Figur \(\(\PageIndex{7}\): Cirkulært, superviklet prokaryotisk DNA. For at gøre det muligt for den store DNA-molekyle at passe ind i bakterien, supervikler et DNA-topoisomeraseenzym kaldet DNA-gyrase kromosomet til et tæt bundt, der danner en komprimeret, superviklet DNA-masse med en diameter på ca. 0,2 µm.
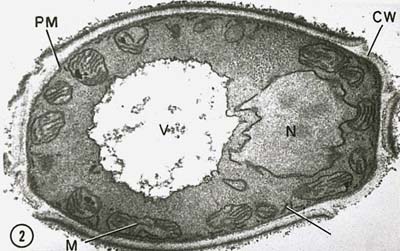
Den prokaryote nukleoid har ingen kernemembran, der omgiver DNA’et, og kernekroppen deler sig ikke ved mitose. Den cytoplasmiske membran spiller en rolle i DNA-separationen under den bakterielle replikation. Da bakterier er haploide (har kun ét kromosom), er der heller ingen meiose.
Det eukaryote genom
Prokaryote og eukaryote celler adskiller sig meget detaljeret fra hinanden både i mængden og organiseringen af deres DNA-molekyler. Eukaryote celler indeholder meget mere DNA end bakterier, og dette DNA er organiseret som flere kromosomer, der er placeret i en kerne.
Kernen i eukaryote celler er omgivet af en kernemembran (Figur \(\PageIndex{7}\)) og indeholder lineære kromosomer, der består af negativt ladet DNA, der er forbundet med positivt ladede basale proteiner kaldet histoner for at danne strukturer, der er kendt som nukleosomer. Nukleosomerne er en del af det, der kaldes kromatin, dvs. det DNA og de proteiner, der udgør kromosomerne. Kernen deler sig ved min mitose, og haploide kønsceller fremstilles af diploide celler ved meiose.
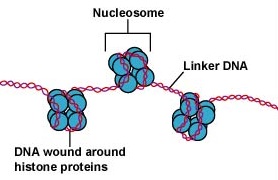
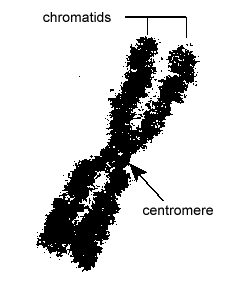
DNA’et i eukaryote celler er pakket på en meget organiseret måde. Det består af en grundlæggende enhed kaldet et nukleosom, en perleagtig struktur på 11 nm i diameter, der består af 146 basepar af DNA, som er viklet om otte histonmolekyler. Nukleosomerne er forbundet med hinanden af et ca. 60 basepar langt DNA-segment, der kaldes linker-DNA (Figur \(\PageIndex{9}\)). En anden histon, der er forbundet med linker-DNA’et, pakker derefter tilstødende nukleotider sammen til en nukleosomtråd med en diameter på 30 nm. Endelig danner disse pakkerede nukleosometråde store sammenrullede sløjfer, der holdes sammen af ikke-histone scaffoldingproteiner. Disse sammenrullede loops på stilladsproteinerne interagerer for at danne det kondenserede kromatin, der ses i kromosomer under mitose (Figur \(\PageIndex{10}\)).
I de senere år har man fundet ud af, at deoxyribonukleoproteinets strukturelle karakter er medvirkende til, om DNA transskriberes til RNA eller ej. For eksempel kan kemiske ændringer i kromatinet gøre det muligt for dele af det at kondensere eller slappe af. Når et område er kondenseret, kan generne ikke transskriberes. Desuden kan kemiske stoffer fæstne sig til eller fjernes fra de histonproteiner, som DNA’et er viklet om. Vedhæftningen eller fjernelsen af disse kemiske grupper til histonet bestemmer, om genekspressionen i nærheden forstærkes eller undertrykkes.
Epigenomet henviser til en række kemiske forbindelser, der ændrer genomet, typisk ved at tilføje en methylgruppe (CH3) til nukleotidbasen adenin på bestemte steder langs DNA-molekylet. Denne methylering kan til gengæld enten undertrykke eller aktivere transkription af specifikke gener. Ved grundlæggende at tænde eller slukke for generne gør epigenomet det muligt for genomet at interagere med og reagere på cellens omgivelser. Epigenomet kan nedarves ligesom genomet.
Summary
- Desoxyribonukleinsyre (DNA) er et langt, dobbeltstrenget, spiralformet molekyle, der består af byggesten kaldet desoxyribonukleotider.
- Et deoxyribonukleotid består af 3 dele: et molekyle af 5-kulstofsukkeret deoxyribose, en nitrogenbas og en fosfatgruppe.
- Der findes fire nitrogenbaser i DNA: adenin, guanin, cytosin eller thymin. Adenin og guanin er kendt som purinbaser, mens cytosin og thymin er kendt som pyrimidinbaser.
- Deoxyribose er et ringformet 5-kulstofsukker. De 5 kulstofatomer er nummereret i rækkefølge med uret rundt om sukkeret. De første 4 kulstofatomer danner faktisk sukkerets ring, idet 5′-kulstoffet kommer fra 4′-kulstoffet i ringen. Nukleotidets nitrogenbase er bundet til sukkerets 1′-kulstof, og fosfatgruppen er bundet til 5′-kulstoffet.
- Under DNA-syntesen kan enzymet DNA-polymerase kun binde fosfatgruppen i et nyt desoxyribonukleotid til 3′-kulstoffet i et nukleotid, der allerede findes i kæden.
- Under DNA-replikationen fungerer hver moderstreng som skabelon for syntesen af den anden streng ved hjælp af komplementær baseparring.
- Komplementær baseparring henviser til, at DNA-nukleotider med basen adenin kun danner hydrogenbindinger med nukleotider, der har basen thymin (A-T). På samme måde kan nukleotider med basen guanin kun hydrogenbindes med nukleotider med basen cytosin (G-C).
- Mens de to DNA-strenge er komplementære, er de orienteret i modsatrettede retninger i forhold til hinanden. Den ene streng siges at løbe 5′ til 3′; den modsatte DNA-streng løber antiparallelt, eller 3′ til 5′.
- I prokaryote celler er der ingen kernemembran, der omgiver DNA’et. Prokaryote celler mangler mitose og meiose.
- For at gøre det muligt for et så stort makromolekyle at passe ind i bakterien binder histonlignende proteiner sig til DNA’et og opdeler DNA-molekylet i omkring 50 kromosomale domæner og gør det mere kompakt. Herefter vikler et enzym kaldet DNA-gyrase hvert domæne om sig selv og danner en komprimeret, superviklet masse af DNA. En topoisomerase kaldet DNA-gyrase katalyserer den negative supercoiling af det cirkulære DNA, der findes i bakterier. Topoisomerase IV er på den anden side involveret i afslapningen af det superoprullede cirkulære DNA, hvilket muliggør adskillelsen af de indbyrdes forbundne datterkromosomer i slutningen af bakteriel DNA-replikation.
- DNA’et i eukaryote celler er pakket i basale enheder kaldet nukleosomer, en perleformet struktur bestående af DNA viklet omkring otte histonmolekyler. DNA’et er organiseret som flere kromosomer, der er placeret i en kerne omgivet af en kernemembran. Kernen deler sig ved mitose, og kønsceller produceres ved meiose hos eukaryoter, der reproducerer sig seksuelt.
- Den strukturelle karakter af deoxyribonukleoproteinet er medvirkende til, om DNA transskriberes til RNA eller ej. For eksempel kan kemiske ændringer af kromatinet gøre det muligt for dele af det at kondensere eller slappe af. Når et område er kondenseret, kan generne ikke transskriberes. Desuden kan kemiske stoffer fæstne sig til eller fjernes fra de histonproteiner, som DNA’et er viklet om. Vedhæftningen eller fjernelsen af disse kemiske grupper til histonet bestemmer, om genekspressionen i nærheden forstærkes eller undertrykkes.
Kontributorer og tilknytninger
-
Dr. Gary Kaiser (COMMUNITY COLLEGE OF BALTIMORE COUNTY, CATONSVILLE CAMPUS)