Lärandemål
- Ange de tre grundläggande delarna av en deoxyribonukleotid.
- Ange vilka kvävebaser som är puriner och vilka som är pyrimidiner.
- Definiera komplementär basparning.
- Gör upp varför DNA endast kan syntetiseras i en 5′- till 3′-riktning.
- Genomför den prokaryotiska nukleoiden med den eukaryotiska kärnan med avseende på följande:
- antal kromosomer
- linjära eller cirkulära kromosomer
- närvaro eller avsaknad av kärnmembran
- närvaro eller avsaknad av nukleosomer
- närvaro eller avsaknad av mitos
- närvaro eller avsaknad av meios
DNA är en lång, dubbelsträngad, spiralformad molekyl som består av byggstenar som kallas desoxyribonukleotider. Varje desoxyribonukleotid består av tre delar: en molekyl av det 5-koliga sockret desoxyribose, en kvävebas och en fosfatgrupp (figur \(\PageIndex{1}\)).
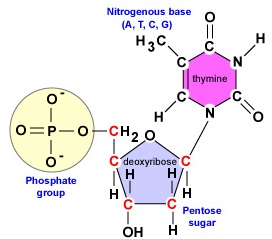
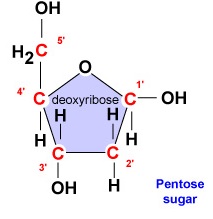
- Deoxyribose. Deoxyribose är ett ringformigt 5-kolssocker (figur \(\PageIndex{2}\)). De 5 kolvätena är numrerade i tur och ordning medurs runt sockret. De fyra första kolvätena bildar faktiskt sockrets ring, där det 5′ kolet kommer från det 4′ kolet i ringen. Nukleotikens kvävebas är bunden till sockrets 1′-kol och fosfatgruppen är bunden till 5′-kolet. Under DNA-syntesen binds fosfatgruppen i en ny desoxyribonukleotid kovalent av enzymet DNA-polymeras till 3′-kolet i en nukleotid som redan finns i kedjan.
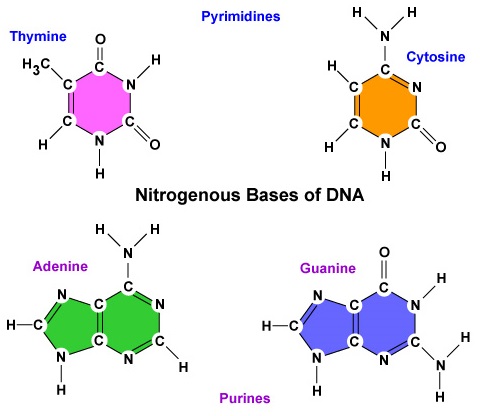
- En kvävebaserad bas. Det finns fyra kvävebaser i DNA: adenin, guanin, cytosin eller tymin. Adenin och guanin kallas purinbaser medan cytosin och tymin kallas pyrimidinbaser (figur \(\PageIndex{3}\)).
- En fosfatgrupp (figur \(\(\PageIndex{4}\)).
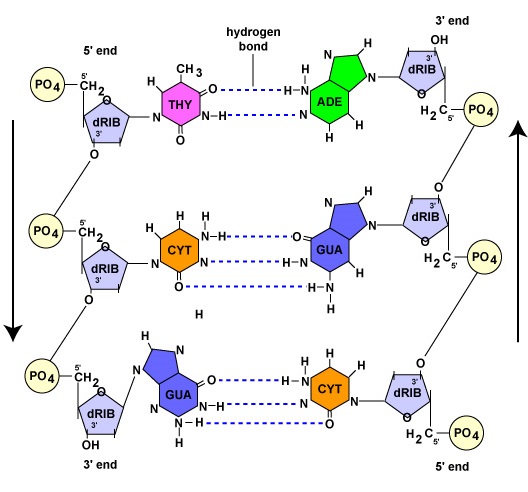
Som vi kommer att se i nästa avsnitt fungerar varje föräldrasträng, under DNA-replikationen, som en mall för syntesen av den andra strängen genom komplementär basparning. Komplementär basparning innebär att DNA-nukleotider med basen adenin endast bildar vätebindningar med nukleotider som har basen tymin (A-T). På samma sätt kan nukleotider med basen guanin vätebindas endast med nukleotider med basen cytosin (G-C). (När det gäller RNA-nukleotider bildar adeninnukleotider, som vi kommer att se senare, vätebindningar med nukleotider med basen uracil eftersom tymin inte finns i RNA). Som ett resultat av denna bindning antar DNA sin spiralform. Därför sägs de två DNA-strängarna vara komplementära. När den ena strängen har en nukleotid som innehåller adenin kommer den motsatta strängen alltid att ha en tymin-nukleotid; när den ena strängen har en nukleotid som innehåller guanin kommer den motsatta strängen alltid att ha en cytosin-nukleotid (figur \(\PageIndex{1}\)).
De två DNA-strängarna är komplementära, men de är orienterade i motsatta riktningar till varandra. Den ena strängen sägs löpa 5′ till 3′; den motsatta DNA-strängen löper antiparallellt, eller 3′ till 5′ (figur \(\PageIndex{1}\)).
Vi kommer nu att kortfattat jämföra prokaryotiska cellers arvsmassa med eukaryotiska cellers arvsmassa.
Det prokaryotiska (bakteriella) genomet
Det område i en bakterie där kromosomen kan ses med ett elektronmikroskop kallas nukleoid. Kromosomen hos de flesta prokaryoter är typiskt sett en enda lång molekyl av dubbelsträngat, spiralformigt, superlindat DNA som bildar en fysisk och genetisk cirkel. Kromosomen är i allmänhet cirka 1000 µm lång och innehåller ofta cirka 4000 gener (figur \(\PageIndex{8}\)). Escherichia coli, som är 2-3 µm lång, har en kromosom som är cirka 1400 µm lång. För att en så stor makromolekyl ska få plats i bakterien binder histonliknande proteiner till DNA:t och delar upp DNA-molekylen i cirka 50 kromosomala domäner, vilket gör den mer kompakt. Ett DNA-topoisomerasenzym som kallas DNA-gyrase spolar sedan upp kromosomen till en tät bunt och bildar en komprimerad, superlindad DNA-massa med en diameter på cirka 0,2 µm.
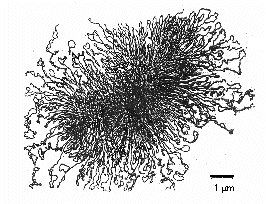
Bakteriella enzymer som kallas DNA-topoisomeraser är viktiga för avveckling, replikation och omspolning av det cirkulära, superrullade bakterie-DNA:t (Figur \(\PageIndex{7}\)). De är också viktiga vid transkription av DNA till RNA, vid DNA-reparation och vid genetisk rekombination i bakterier.
Figur \(\PageIndex{7}\): Cirkulärt, superlindat prokaryotiskt DNA. För att den stora DNA-molekylen ska kunna rymmas i bakterien, superlindas kromosomen av ett DNA-topoisomerasenzym som kallas DNA-gyrase till en tät bunt som bildar en komprimerad, superlindad DNA-massa med en diameter på cirka 0,2 µm.
Den prokaryotiska nukleoiden har inget kärnmembran som omger DNA:t, och kärnkroppen delar sig inte genom mitos. Det cytoplasmatiska membranet spelar en roll vid DNA-separation under bakteriell replikation. Eftersom bakterier är haploida (har bara en kromosom) förekommer inte heller någon meios.
Den eukaryotiska arvsmassan
Prokaryotiska och eukaryotiska celler skiljer sig åt på ett mycket detaljerat sätt både när det gäller mängden och organisationen av deras DNA-molekyler. Eukaryota celler innehåller mycket mer DNA än bakterier, och detta DNA är organiserat som flera kromosomer som är placerade i en kärna.
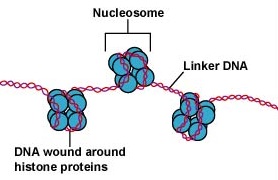
Kärnan i eukaryota celler är omgiven av ett kärnmembran (figur \(\(\PageIndex{7}\))) och innehåller linjära kromosomer som består av negativt laddat DNA som är associerat med positivt laddade basiska proteiner, så kallade histoner, för att bilda strukturer som kallas nukleosomer. Nukleosomerna är en del av det som kallas kromatin, det DNA och de proteiner som utgör kromosomerna. Kärnan delar sig genom mitos och haploida könsceller produceras från diploida celler genom meios.
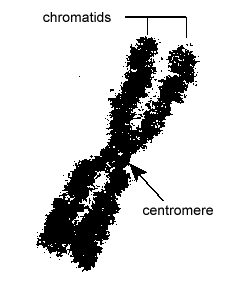
DNA:t i eukaryota celler är förpackat på ett mycket organiserat sätt. Det består av en grundläggande enhet som kallas nukleosom, en pärlliknande struktur med en diameter på 11 nm som består av 146 baspar DNA lindat runt åtta histonmolekyler. Nukleosomerna är sammanlänkade med varandra genom ett cirka 60 baspar långt DNA-segment som kallas länk-DNA (figur \(\PageIndex{9}\)). En annan histon som är associerad med länk-DNA:t paketerar sedan intilliggande nukleotider tillsammans för att bilda en nukleosomtråd med en diameter på 30 nm. Slutligen bildar dessa paketerade nukleosomtrådar stora spiralformade slingor som hålls samman av icke-histoneffektiva scaffoldingproteiner. Dessa spiralformade slingor på de olika ställningsskyddsproteinerna samverkar för att bilda det kondenserade kromatin som ses i kromosomerna under mitos (figur \(\PageIndex{10}\)).
Under senare år har man funnit att deoxyribonukleoproteinets strukturella karaktär bidrar till om DNA transkriberas till RNA eller inte. Till exempel kan kemiska förändringar av kromatinet göra det möjligt för delar av det att kondensera eller slappna av. När en region är kondenserad kan generna inte transkriberas. Dessutom kan kemiska ämnen fästa vid eller avlägsnas från de histonproteiner som DNA:t omsluter. Fastsättningen eller avlägsnandet av dessa kemiska grupper på histonet avgör om det närliggande genuttrycket förstärks eller undertrycks.
Epigenomet avser en mängd olika kemiska föreningar som ändrar genomet vanligtvis genom att lägga till en metylgrupp (CH3) till nukleotidbasen adenin på specifika platser längs DNA-molekylen. Denna metylering kan i sin tur antingen undertrycka eller aktivera transkriptionen av specifika gener. Genom att i princip slå på eller stänga av gener gör epigenomet det möjligt för genomet att interagera med och reagera på cellens miljö. Epigenomet kan ärvas precis som genomet.
Sammanfattning
- Deoxyribonukleinsyra (DNA) är en lång, dubbelsträngad, spiralformad molekyl som består av byggstenar som kallas desoxyribonukleotider.
- En desoxyribonukleotid består av 3 delar: en molekyl av det 5-koliga sockret desoxyribose, en kvävebas och en fosfatgrupp.
- Det finns fyra kvävebaser i DNA: adenin, guanin, cytosin eller tymin. Adenin och guanin är så kallade purinbaser medan cytosin och tymin är så kallade pyrimidinbaser.
- Deoxyribose är ett ringformigt socker med 5 kol. De 5 kolvätena är numrerade i följd medurs runt sockret. De första 4 kolvätena bildar faktiskt sockrets ring med 5′-kolet som kommer från 4′-kolet i ringen. Nukleotikens kvävebas är knuten till sockrets 1′-kol och fosfatgruppen är bunden till 5′-kolet.
- Under DNA-syntesen kan enzymet DNA-polymeras endast knyta fosfatgruppen i en ny desoxyribonukleotid till 3′-kolet i en nukleotid som redan finns i kedjan.
- Under DNA-replikationen fungerar varje föräldrasträng som en mall för syntesen av den andra strängen genom komplementär basparning.
- Komplementär basparning avser DNA-nukleotider med basen adenin som endast bildar vätebindningar med nukleotider som har basen tymin (A-T). På samma sätt kan nukleotider med basen guanin endast vätebindas med nukleotider med basen cytosin (G-C).
- Medans de två DNA-strängarna är komplementära är de orienterade i motsatt riktning till varandra. Den ena strängen sägs löpa 5′ till 3′; den motsatta DNA-strängen löper antiparallellt, eller 3′ till 5′.
- I prokaryota celler finns det inget kärnmembran som omger DNA. Prokaryota celler saknar mitos och meios.
- För att en så här stor makromolekyl ska få plats i bakterien binder histonliknande proteiner till DNA:t, vilket segregerar DNA-molekylen i cirka 50 kromosomala domäner och gör den mer kompakt. Ett enzym som kallas DNA-gyrase spolar sedan varje domän runt sig själv och bildar en komprimerad, superlindad DNA-massa. Ett topoisomeras som kallas DNA-gyrase katalyserar den negativa supercoilingen av det cirkulära DNA som finns i bakterier. Topoisomeras IV är å andra sidan inblandat i relaxationen av det superlindade cirkulära DNA:t, vilket gör det möjligt att separera de sammankopplade dotterkromosomerna i slutet av bakteriell DNA-replikation.
- DNA:t i eukaryota celler är paketerat i grundläggande enheter som kallas nukleosomer, en pärlliknande struktur som består av DNA som är lindat runt åtta histonmolekyler. DNA:t är organiserat som flera kromosomer som ligger i en kärna omgiven av ett kärnmembran. Kärnan delas genom mitos och könsceller produceras genom meios hos eukaryoter som reproducerar sig sexuellt.
- Den strukturella karaktären hos desoxyribonukleoproteinet bidrar till om DNA transkriberas till RNA eller inte. Till exempel kan kemiska förändringar av kromatinet göra det möjligt för delar av det att kondensera eller slappna av. När en region är kondenserad kan gener inte transkriberas. Dessutom kan kemiska ämnen fästa vid eller avlägsnas från de histonproteiner som DNA:t omsluter. Hur dessa kemiska grupper fäster eller avlägsnas från histonerna avgör om genuttrycket i närheten förstärks eller undertrycks.
Kontributer och tillskrivningar
-
Dr. Gary Kaiser (COMMUNITY COLLEGE OF BALTIMORE COUNTY, CATONSVILLE CAMPUS)