Obiettivi di apprendimento
- Dichiarate le tre parti fondamentali di un deossiribonucleotide.
- Dimostrare quali basi azotate sono purine e quali pirimidine.
- Definire l’accoppiamento di basi complementari.
- Dimmi perché il DNA può essere sintetizzato solo in direzione 5′ a 3′.
- Confrontare il nucleoide procariotico con il nucleo eucariotico in termini di quanto segue:
- numero di cromosomi
- cromosomi lineari o circolari
- presenza o assenza di una membrana nucleare
- presenza o assenza di nucleosomi
- presenza o assenza di mitosi
- presenza o assenza di meiosi
Il DNA è una lunga a doppio filamento, molecola elicoidale composta da blocchi di costruzione chiamati deossiribonucleotidi. Ogni deossiribonucleotide è composto da tre parti: una molecola di zucchero deossiribosio a 5 carboni, una base azotata e un gruppo fosfato (Figura \(\PageIndex{1}).
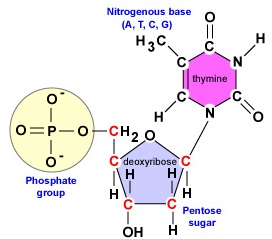
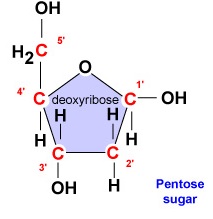
- Deossiribosio. Il desossiribosio è uno zucchero ad anello a 5 carboni (Figura \PageIndex{2}\). I 5 carboni sono numerati sequenzialmente in senso orario intorno allo zucchero. I primi 4 carboni formano effettivamente l’anello dello zucchero con il 5′ carbonio che si stacca dal 4′ carbonio dell’anello. La base azotata del nucleotide è attaccata al carbonio 1′ dello zucchero e il gruppo fosfato è legato al carbonio 5′. Durante la sintesi del DNA, il gruppo fosfato di un nuovo deossiribonucleotide è legato covalentemente dall’enzima DNA polimerasi al carbonio 3′ di un nucleotide già presente nella catena.
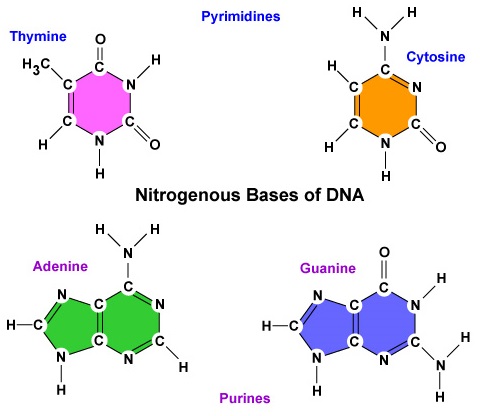
- Una base azotata. Ci sono quattro basi azotate nel DNA: adenina, guanina, citosina o timina. L’adenina e la guanina sono conosciute come basi puriniche, mentre la citosina e la timina sono conosciute come basi pirimidiniche (Figura \PageIndex{3}).
- Un gruppo fosfato (Figura \(\PageIndex{4}).
Per sintetizzare le due catene di deossiribonucleotidi durante la replicazione del DNA, gli enzimi DNA polimerasi coinvolti sono solo in grado di unire il gruppo fosfato al carbonio 5′ di un nuovo nucleotide al gruppo idrossile (OH) del carbonio 3′ di un nucleotide (Figura \(\PageIndex{2}) già presente nella catena. Il legame covalente che unisce i nucleotidi è chiamato legame fosfodiestere. Ogni filamento di DNA ha una cosiddetta estremità 5′ e una estremità 3′. Questo significa che un’estremità di ogni filamento di DNA, chiamata estremità 5′, avrà sempre un gruppo fosfato attaccato al carbonio 5′ del suo deossiribonucleotide terminale (Figura \(\PageIndex{5})). L’altra estremità di quel filamento, chiamata estremità 3′, avrà sempre un idrossile (OH) sul carbonio 3′ del suo deossiribonucleotide terminale.
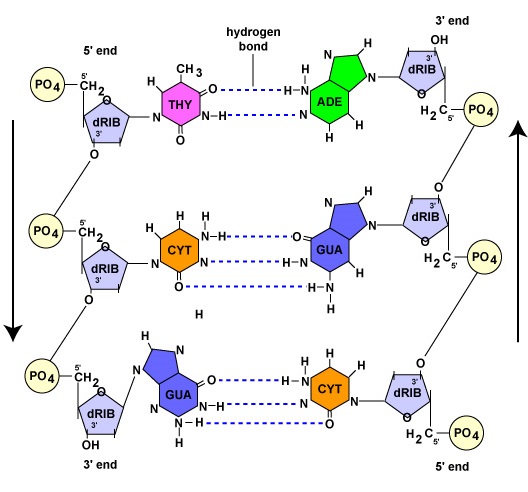
Come si vedrà nella prossima sezione, ogni filamento genitore, durante la replicazione del DNA, agisce come modello per la sintesi dell’altro filamento attraverso l’accoppiamento complementare di basi. L’accoppiamento di base complementare si riferisce ai nucleotidi del DNA con la base adenina che formano solo legami idrogeno con i nucleotidi che hanno la base timina (A-T). Allo stesso modo, i nucleotidi con la base guanina possono legarsi a idrogeno solo con i nucleotidi che hanno la base citosina (G-C). (Nel caso dei nucleotidi dell’RNA, come si vedrà più avanti, i nucleotidi di adenina formano legami a idrogeno con i nucleotidi di base uracile, poiché la timina non si trova nell’RNA). Come risultato di questo legame, il DNA assume la sua forma elicoidale. Pertanto, i due filamenti di DNA sono detti complementari. Ovunque un filamento abbia un nucleotide contenente adenina, il filamento opposto avrà sempre un nucleotide di timina; ovunque ci sia un nucleotide contenente guanina, il filamento opposto avrà sempre un nucleotide di citosina (Figura \(\PageIndex{1})).
Mentre i due filamenti di DNA sono complementari, sono orientati in direzioni opposte l’uno all’altro. Si dice che un filamento corre da 5′ a 3′; il filamento di DNA opposto corre antiparallelo, o da 3′ a 5′ (Figura \PageIndex{1}}).
Confronteremo ora brevemente il genoma delle cellule procariote con quello delle cellule eucariote.
Il genoma procariotico (batterico)
L’area all’interno di un batterio dove il cromosoma può essere visto con un microscopio elettronico è chiamato nucleoide. Il cromosoma della maggior parte dei procarioti è tipicamente una lunga, singola molecola di DNA a doppio filamento, elicoidale e superavvolto che forma un cerchio fisico e genetico. Il cromosoma è generalmente lungo circa 1000 µm e spesso contiene circa 4000 geni (Figura \(\PageIndex{8})). Escherichia coli, che è lungo 2-3 µm, ha un cromosoma lungo circa 1400 µm. Per permettere a una macromolecola così grande di adattarsi al batterio, proteine simili agli istoni si legano al DNA, segregando la molecola di DNA in circa 50 domini cromosomici e rendendola più compatta. Un enzima topoisomerasi del DNA, chiamato DNA gyrase, avvolge il cromosoma in un fascio stretto, formando una massa di DNA compattata e superavvolta di circa 0,2 µm di diametro.
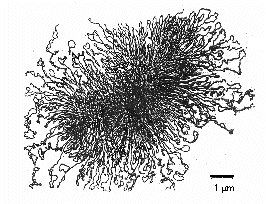
Gli enzimi batterici chiamati topoisomerasi del DNA sono essenziali nello svolgimento, replicazione e riavvolgimento del DNA batterico circolare e superavvolto (Figura \PageIndex{7}). Sono anche essenziali nella trascrizione del DNA in RNA, nella riparazione del DNA e nella ricombinazione genetica nei batteri.
Figura \(\PageIndex{7}): DNA procariotico circolare e superavvolto. Per permettere alla grande molecola di DNA di adattarsi all’interno del batterio, un enzima topoisomerasi del DNA chiamato DNA girasi avvolge il cromosoma in un fascio stretto formando una massa compattata e superavvolta di DNA di circa 0,2 µm di diametro.
Il nucleoide procariotico non ha membrana nucleare che circonda il DNA e il corpo nucleare non si divide per mitosi. La membrana citoplasmatica gioca un ruolo nella separazione del DNA durante la replicazione batterica. Poiché i batteri sono aploidi (hanno un solo cromosoma), non c’è nemmeno la meiosi.
Il genoma eucariotico
Le cellule procariotiche ed eucariotiche differiscono molto sia nella quantità che nell’organizzazione delle loro molecole di DNA. Le cellule eucariotiche contengono molto più DNA rispetto ai batteri, e questo DNA è organizzato come cromosomi multipli situati all’interno di un nucleo.
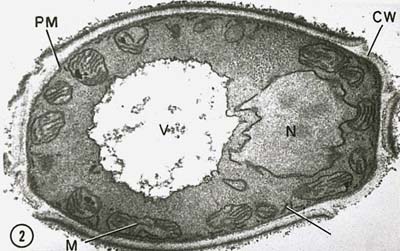
Il nucleo nelle cellule eucariotiche è circondato da una membrana nucleare (Figura \(\PageIndex{7})) e contiene cromosomi lineari composti da DNA a carica negativa associati a proteine di base a carica positiva chiamate istoni per formare strutture note come nucleosomi. I nucleosomi fanno parte di quella che si chiama cromatina, il DNA e le proteine che compongono i cromosomi. Il nucleo si divide per mitosi e le cellule sessuali aploidi sono prodotte da cellule diploidi per meiosi.
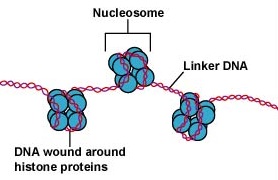
Il DNA nelle cellule eucariotiche è confezionato in modo altamente organizzato. Consiste in un’unità di base chiamata nucleosoma, una struttura simile a una perlina di 11 nm di diametro che consiste di 146 coppie di basi di DNA avvolte intorno a otto molecole di istone. I nucleosomi sono collegati tra loro da un segmento di DNA lungo circa 60 paia di basi chiamato DNA linker (Figura \PageIndex{9}}). Un altro istone associato al DNA linker impacchetta i nucleotidi adiacenti per formare un filo di nucleosoma di 30 nm di diametro. Infine, questi fili di nucleosomi impacchettati formano grandi anelli arrotolati che sono tenuti insieme da proteine di impalcatura non istone. Questi anelli arrotolati sulle proteine di impalcatura interagiscono per formare la cromatina condensata vista nei cromosomi durante la mitosi (Figura \PageIndex{10}}).
Negli ultimi anni si è scoperto che la natura strutturale della desossiribonucleoproteina contribuisce a far sì che il DNA sia trascritto o meno in RNA. Per esempio, i cambiamenti chimici alla cromatina possono permettere a porzioni di essa di condensarsi o rilassarsi. Quando una regione è condensata, i geni non possono essere trascritti. Inoltre, le sostanze chimiche possono attaccarsi o essere rimosse dalle proteine istone intorno alle quali il DNA si avvolge. L’attaccamento o la rimozione di questi gruppi chimici all’istone determina se l’espressione del gene vicino è amplificata o repressa.
L’epigenoma si riferisce ad una varietà di composti chimici che modificano il genoma tipicamente aggiungendo un gruppo metile (CH3) alla base nucleotidica adenina in punti specifici lungo la molecola del DNA. Questa metilazione può, a sua volta, reprimere o attivare la trascrizione di geni specifici. Accendendo o spegnendo i geni, l’epigenoma permette al genoma di interagire e rispondere all’ambiente della cellula. L’epigenoma può essere ereditato proprio come il genoma.
Sommario
- L’acido desossiribonucleico (DNA) è una lunga molecola elicoidale a doppio filamento, composta da elementi chiamati deossiribonucleotidi.
- Un deossiribonucleotide è composto da 3 parti: una molecola di zucchero deossiribosio a 5 carboni, una base azotata e un gruppo fosfato.
- Ci sono quattro basi azotate nel DNA: adenina, guanina, citosina o timina. L’adenina e la guanina sono conosciute come basi puriniche, mentre la citosina e la timina sono conosciute come basi pirimidiniche.
- Il desossiribosio è uno zucchero a 5 carboni ad anello. I 5 carboni sono numerati sequenzialmente in senso orario intorno allo zucchero. I primi 4 carboni formano effettivamente l’anello dello zucchero con il 5′ carbonio che si stacca dal 4′ carbonio dell’anello. La base azotata del nucleotide è attaccata al carbonio 1′ dello zucchero e il gruppo fosfato è legato al carbonio 5′.
- Nella sintesi del DNA, l’enzima DNA polimerasi può attaccare il gruppo fosfato di un nuovo deossiribonucleotide solo al carbonio 3′ di un nucleotide già presente nella catena.
- Durante la replicazione del DNA, ogni filamento genitore funge da modello per la sintesi dell’altro filamento per mezzo dell’appaiamento di basi complementari.
- L’appaiamento di basi complementari si riferisce ai nucleotidi del DNA con la base adenina che formano solo legami idrogeno con nucleotidi che hanno la base timina (A-T). Allo stesso modo, i nucleotidi con la base guanina possono legarsi a idrogeno solo con i nucleotidi che hanno la base citosina (G-C).
- Mentre i due filamenti di DNA sono complementari, essi sono orientati in direzioni opposte l’uno all’altro. Si dice che un filamento corre da 5′ a 3′; il filamento di DNA opposto corre antiparallelo, o da 3′ a 5′.
- Nelle cellule procariotiche non c’è una membrana nucleare che circonda il DNA. Le cellule procariotiche mancano di mitosi e meiosi.
- Per permettere a una macromolecola così grande di adattarsi al batterio, proteine simili agli istoni si legano al DNA, segregando la molecola di DNA in circa 50 domini cromosomici e rendendola più compatta. Poi un enzima chiamato DNA gyrase avvolge ogni dominio su se stesso formando una massa di DNA compattata e superavvolta. Una topoisomerasi chiamata DNA gyrase catalizza il superavvolgimento negativo del DNA circolare che si trova nei batteri. La topoisomerasi IV, d’altra parte, è coinvolta nel rilassamento del DNA circolare superavvolto, consentendo la separazione dei cromosomi figli interconnessi alla fine della replicazione del DNA batterico.
- Il DNA nelle cellule eucariotiche è impacchettato in unità di base chiamate nucleosomi, una struttura simile a perline che consiste di DNA avvolto intorno a otto molecole di istone. Il DNA è organizzato come cromosomi multipli situati all’interno di un nucleo circondato da una membrana nucleare. Il nucleo si divide per mitosi e i gameti sono prodotti dalla meiosi negli eucarioti che si riproducono sessualmente.
- La natura strutturale della desossiribonucleoproteina contribuisce a far sì che il DNA sia trascritto o meno in RNA. Per esempio, i cambiamenti chimici alla cromatina possono permettere a porzioni di essa di condensarsi o rilassarsi. Quando una regione è condensata, i geni non possono essere trascritti. Inoltre, le sostanze chimiche possono attaccarsi o essere rimosse dalle proteine istone intorno alle quali il DNA si avvolge. L’attaccamento o la rimozione di questi gruppi chimici all’istone determina se l’espressione del gene vicino è amplificata o repressa.
Contribuenti e attribuzioni
-
Dr. Gary Kaiser (COMMUNITY COLLEGE OF BALTIMORE COUNTY, CATONSVILLE CAMPUS)