Tanulási célok
- Mondja meg a dezoxiribonukleotid három alapvető részét.
- Mondja meg, hogy mely nitrogénbázisok purinok és melyek pirimidinek.
- Meghatározza a komplementer bázispárosítást.
- Mondja meg, hogy miért csak 5′ 3′ irányban lehet DNS-t szintetizálni.
- Hasonlítsa össze a prokarióta nukleoidot az eukarióta sejtmaggal az alábbiak szempontjából:
- a kromoszómák száma
- lineáris vagy körkörös kromoszómák
- magmembrán jelenléte vagy hiánya
- nukleoszómák jelenléte vagy hiánya
- mitózis jelenléte vagy hiánya
- meiózis jelenléte vagy hiánya
A DNS egy hosszú, kettős szálú, spirális molekula, amely dezoxiribonukleotidoknak nevezett építőelemekből áll. Minden dezoxiribonukleotid három részből áll: az 5 szénatomos dezoxiribóz cukor molekulájából, egy nitrogén bázisból és egy foszfátcsoportból (\(\PageIndex{1}\) ábra).
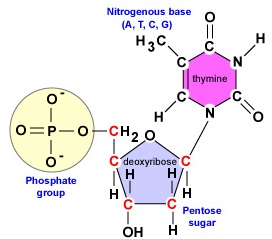
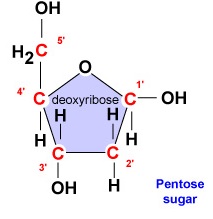
- Dezoxiribóz. A dezoxiribóz egy gyűrűs 5 szénatomos cukor (\(\PageIndex{2}\ ábra)). Az 5 szénatomot az óramutató járásával megegyezően sorszámozzuk a cukor körül. Az első 4 szénatom valójában a cukor gyűrűjét alkotja, az 5′ szénatom a gyűrű 4′ szénatomjáról származik. A nukleotid nitrogénbázisa a cukor 1′ szénatomjához kapcsolódik, a foszfátcsoport pedig az 5′ szénatomhoz. A DNS-szintézis során egy új dezoxiribonukleotid foszfátcsoportját a DNS-polimeráz enzim kovalensen hozzákapcsolja a láncban már meglévő nukleotid 3′ szénatomjához.
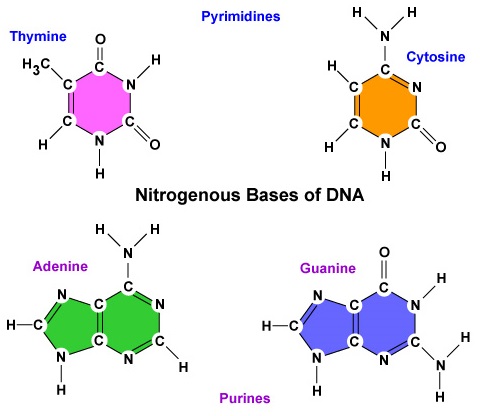
- A nitrogénbázis. A DNS-ben négy nitrogén bázis található: adenin, guanin, citozin vagy timin. Az adenint és a guanint purinbázisoknak, míg a citozint és a timint pirimidinbázisoknak nevezzük (\(\PageIndex{3}\ ábra)).
- Egy foszfátcsoport (\(\PageIndex{4}\) ábra).
A DNS-replikáció során a dezoxiribonukleotidok két láncának szintéziséhez a DNS-polimeráz enzimek csak arra képesek, hogy egy új nukleotid 5′ szénatomjánál lévő foszfátcsoportot egy már a láncban lévő nukleotid (\(\PageIndex{2}\) ábra) 3′ szénatomjának hidroxil (OH) csoportjához kapcsolják. A nukleotidokat összekötő kovalens kötést foszfodiészterkötésnek nevezzük. Minden DNS-szálnak van egy úgynevezett 5′ és egy 3′ vége. Ez azt jelenti, hogy minden DNS-szál egyik végéhez, az 5′ véghez mindig egy foszfátcsoport kapcsolódik a terminális dezoxiribonukleotid 5′ szénatomjához (\(\PageIndex{5}\) ábra). A szál másik vége, a 3′ vég, mindig hidroxilcsoportot (OH) tartalmaz a terminális dezoxiribonukleotid 3′ szénatomján.
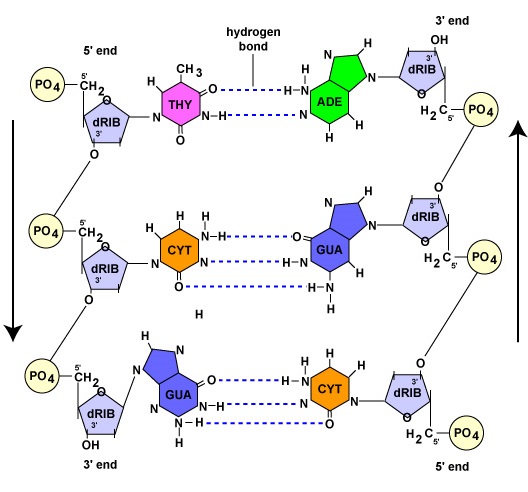
Amint azt a következő részben látni fogjuk, a DNS-replikáció során mindkét szülői szál a komplementer bázispárosodás révén templátként szolgál a másik szál szintéziséhez. A komplementer bázispárosodás arra utal, hogy az adenin bázissal rendelkező DNS-nukleotidok csak a timin bázissal (A-T) rendelkező nukleotidokkal alkotnak hidrogénkötést. Hasonlóképpen, a guanin bázissal rendelkező nukleotidok csak a citozin bázissal (G-C) rendelkező nukleotidokkal képesek hidrogénkötésre. (Az RNS-nukleotidok esetében, mint később látni fogjuk, az adenin nukleotidok hidrogénkötést képeznek az uracil bázissal rendelkező nukleotidokkal, mivel a timin nem található meg az RNS-ben.) Az RNS-nukleotidok hidrogénkötést képeznek az uracil bázissal rendelkező nukleotidokkal. E kötés eredményeként a DNS felveszi spirális alakját. Ezért mondjuk, hogy a DNS két szála komplementer. Ahol az egyik szál adenintartalmú nukleotiddal rendelkezik, ott a másik szál mindig timinnel; ahol guanintartalmú nukleotid van, ott a másik szál mindig citozinnal rendelkezik (\(\PageIndex{1}\) ábra).
Míg a DNS két szála komplementer, addig egymással ellentétes irányban orientálódnak. Az egyik szálról azt mondjuk, hogy 5′-től 3′-ig tart; az ellentétes DNS-szál pedig antiparallel, azaz 3′-től 5′-ig tart (\(\PageIndex{1}\ ábra)).
A következőkben röviden összehasonlítjuk a prokarióta sejtek genomját az eukarióta sejtekével.
A prokarióta (baktérium) genom
A baktériumon belüli területet, ahol a kromoszóma elektronmikroszkóppal látható, nukleoidnak nevezzük. A legtöbb prokarióta kromoszómája jellemzően egy hosszú, egyetlen molekula kettős szálú, spirális, szupertekercselt DNS, amely egy fizikai és genetikai kört alkot. A kromoszóma általában körülbelül 1000 µm hosszú, és gyakran körülbelül 4000 gént tartalmaz (\(\PageIndex{8}\) ábra). A 2-3 µm hosszúságú Escherichia coli kromoszómája körülbelül 1400 µm hosszú. Ahhoz, hogy egy ekkora makromolekula elférjen a baktériumban, hisztonszerű fehérjék kötődnek a DNS-hez, amelyek a DNS-molekulát mintegy 50 kromoszómális doménre osztják szét, és kompaktabbá teszik. A DNS topoizomeráz enzim, a DNS gyráz ezután a kromoszómát szoros köteggé tekeri, és egy tömörített, szupertekercselt DNS-tömeget képez, amelynek átmérője körülbelül 0,2 µm.
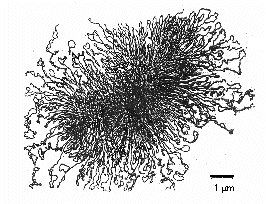
A DNS topoizomerázoknak nevezett bakteriális enzimek nélkülözhetetlenek a körkörös, szupertekercselt bakteriális DNS kitekerésében, replikációjában és visszatekerésében (\(\(\PageIndex{7}\) ábra). Elengedhetetlenek továbbá a DNS RNS-é történő átírásában, a DNS-javításban és a baktériumok genetikai rekombinációjában.
\(\PageIndex{7}\) ábra: Körkörös, szupertekercses prokarióta DNS. Annak érdekében, hogy a nagy DNS-molekula elférjen a baktériumban, a DNS topoizomeráz enzim, a DNS gyráz szuper tekercseli a kromoszómát egy szoros köteggé, amely egy tömör, szuper tekercselt DNS tömeget alkot, körülbelül 0,2 µm átmérővel.
A prokarióta nukleoidot nem veszi körül magmembrán, és a magtest nem osztódik mitózis útján. A citoplazmamembrán a DNS szétválasztásában játszik szerepet a bakteriális replikáció során. Mivel a baktériumok haploidok (csak egy kromoszómával rendelkeznek), nincs meiózis sem.
Az eukarióta genom
A prokarióta és az eukarióta sejtek mind a DNS-molekuláik mennyiségében, mind a szervezetükben nagyban különböznek egymástól. Az eukarióta sejtek sokkal több DNS-t tartalmaznak, mint a baktériumok, és ez a DNS a sejtmagban elhelyezkedő több kromoszómába szerveződik.
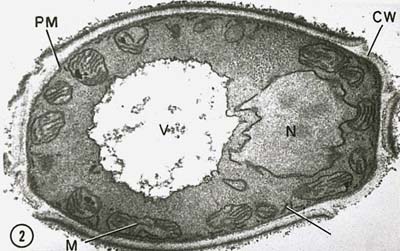
Az eukarióta sejtek sejtmagját magmembrán veszi körül (\(\PageIndex{7}\ ábra)), és negatív töltésű DNS-ből álló lineáris kromoszómákat tartalmaz, amelyek pozitív töltésű alapfehérjékhez, a hisztonokhoz kapcsolódva nukleoszómáknak nevezett szerkezeteket alkotnak. A nukleoszómák részét képezik az úgynevezett kromatin, a kromoszómákat alkotó DNS és fehérjék. A sejtmag osztódik a mitózissal, és a diploid sejtekből a meiózissal haploid ivarsejtek keletkeznek.
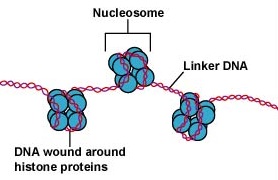
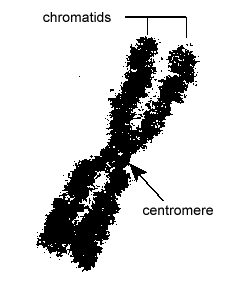
A DNS az eukarióta sejtekben rendkívül szervezett módon van becsomagolva. Egy nukleoszómának nevezett alapegységből áll, egy 11 nm átmérőjű gyöngyszerű struktúrából, amely 146 bázispár DNS-ből áll, nyolc hisztonmolekula köré tekeredve. A nukleoszómákat egy körülbelül 60 bázispár hosszú DNS-szegmens, az úgynevezett linker-DNS köti össze (\(\PageIndex{9}\) ábra). A linker-DNS-hez kapcsolódó másik hiszton ezután a szomszédos nukleotidokat egy 30 nm átmérőjű nukleoszóma-szálat alkotva csomagolja össze. Végül ezek a csomagolt nukleoszóma szálak nagy tekercselt hurkokat alkotnak, amelyeket nem hisztonból álló állványozó fehérjék tartanak össze. Ezek a tekercselt hurkok az állványozó fehérjékkel kölcsönhatásba lépve alkotják a mitózis során a kromoszómákban látható kondenzált kromatint (\(\PageIndex{10}\) ábra).
Az utóbbi években kiderült, hogy a dezoxiribonukleoprotein szerkezeti jellege hozzájárul ahhoz, hogy a DNS-t RNS-vé írják-e át vagy sem. A kromatin kémiai változásai például lehetővé tehetik, hogy a kromatin egyes részei kondenzálódjanak vagy lazuljanak. Ha egy régió kondenzálódik, a gének nem tudnak átíródni. Ezenkívül kémiai anyagok kapcsolódhatnak a DNS-t körülvevő hisztonfehérjékhez vagy eltávolodhatnak azokról. Ezeknek a kémiai csoportoknak a hisztonhoz való kapcsolódása vagy eltávolítása határozza meg, hogy a közeli génexpresszió felerősödik vagy elnyomódik.
Az epigenom olyan kémiai vegyületek sokaságára utal, amelyek a genomot módosítják, jellemzően úgy, hogy a DNS-molekula mentén meghatározott helyeken metil (CH3) csoportot adnak a nukleotid bázis adeninjéhez. Ez a metiláció viszont elnyomhatja vagy aktiválhatja bizonyos gének átírását. Alapvetően a gének be- vagy kikapcsolásával az epigenom lehetővé teszi, hogy a genom kölcsönhatásba lépjen a sejt környezetével és reagáljon arra. Az epigenom a genomhoz hasonlóan öröklődhet.
Összefoglaló
- A dezoxiribonukleinsav (DNS) egy hosszú, kettős szálú, spirális molekula, amely dezoxiribonukleotidoknak nevezett építőelemekből áll.
- A dezoxiribonukleotid 3 részből áll: az 5 szénatomos dezoxiribóz cukor molekulájából, egy nitrogén bázisból és egy foszfátcsoportból.
- A DNS-ben négy nitrogén bázis található: adenin, guanin, citozin vagy timin. Az adenint és a guanint purinbázisoknak, míg a citozint és a timint pirimidinbázisoknak nevezzük.
- A dezoxiribóz egy gyűrűs 5 szénatomos cukor. Az 5 szénatomot az óramutató járásával megegyezően sorszámozzák a cukor körül. Az első 4 szénatom valójában a cukor gyűrűjét alkotja, az 5′ szénatom a gyűrű 4′ szénatomjáról származik. A nukleotid nitrogénbázisa a cukor 1′ szénatomjához kapcsolódik, a foszfátcsoport pedig az 5′ szénatomhoz.
- A DNS-szintézis során a DNS-polimeráz enzim csak egy már a láncban lévő nukleotid 3′ szénatomjához tudja egy új dezoxiribonukleotid foszfátcsoportját kötni.
- A DNS-replikáció során az egyes szülői szálak komplementer bázispárosítás révén sablonként szolgálnak a másik szál szintéziséhez.
- A komplementer bázispárosítás arra utal, hogy az adenin bázissal rendelkező DNS-nukleotidok csak a timin bázissal (A-T) rendelkező nukleotidokkal alkotnak hidrogénkötést. Hasonlóképpen, a guanin bázissal rendelkező nukleotidok csak a citozin bázissal (G-C) rendelkező nukleotidokkal képesek hidrogénkötésre.
- Míg a DNS két szála komplementer, addig egymással ellentétes irányban orientálódnak. Az egyik szálról azt mondjuk, hogy 5′-től 3′-ig tart; az ellentétes DNS-szál antiparalel, vagyis 3′-től 5′-ig tart.
- A prokarióta sejtekben nincs a DNS-t körülvevő magmembrán. A prokarióta sejtekből hiányzik a mitózis és a meiózis.
- Azért, hogy egy ekkora makromolekula elférjen a baktériumban, hisztonszerű fehérjék kötődnek a DNS-hez, mintegy 50 kromoszómális tartományra osztva és tömörebbé téve a DNS-molekulát. Ezután a DNS-giráz nevű enzim minden egyes tartományt maga köré tekercsel, és így egy tömörített, szupertekercselt DNS-tömeget alkot. A DNS-giráz nevű topoizomeráz katalizálja a baktériumokban található cirkuláris DNS negatív szupercoilingját. A IV. topoizomeráz viszont részt vesz a szupertekercselt cirkuláris DNS lazításában, lehetővé téve az összekapcsolt leánykromoszómák szétválasztását a bakteriális DNS-replikáció végén.
- A DNS az eukarióta sejtekben nukleoszómáknak nevezett alapegységekbe van csomagolva, egy gyöngyszerű szerkezetbe, amely nyolc hisztonmolekula köré tekert DNS-ből áll. A DNS többszörös kromoszómákba szerveződik, amelyek a magmembránnal körülvett sejtmagban helyezkednek el. A sejtmag mitózis útján osztódik, és az ivarosan szaporodó eukariótákban az ivarsejtek meiózissal jönnek létre.
- A dezoxiribonukleoprotein szerkezeti jellege hozzájárul ahhoz, hogy a DNS RNS-é íródik-e át vagy sem. A kromatin kémiai változásai például lehetővé tehetik, hogy a kromatin egyes részei kondenzálódjanak vagy lazuljanak. Ha egy régió kondenzálódik, a gének nem íródnak át. Ezenkívül kémiai anyagok kapcsolódhatnak a DNS-t körülvevő hisztonfehérjékhez vagy eltávolodhatnak azokról. Ezeknek a kémiai csoportoknak a hisztonhoz való kapcsolódása vagy eltávolítása határozza meg, hogy a közeli génexpresszió felerősödik vagy elnyomódik.
Megosztók és hozzászólások
-
Dr. Gary Kaiser (COMMUNITY COLLEGE OF BALTIMORE COUNLEY, CATONSVILLE CAMPUS)