Oppimistavoitteet
- Nimeä deoksiribonukleotidin kolme perusosaa.
- Neuvo, mitkä typpiemäkset ovat puriineja ja mitkä pyrimidiiniä.
- Määrittele komplementaarinen emäspariutuminen.
- Kerro, miksi DNA:ta voidaan syntetisoida vain 5′ 3′ -suunnassa.
- Vertaile prokaryoottista nukleoidia eukaryoottiseen ytimeen seuraavien seikkojen suhteen:
- kromosomien lukumäärä
- lineaariset vai pyöreät kromosomit
- ydinkalvon esiintyminen tai puuttuminen
- nukleosomien esiintyminen tai puuttuminen
- mitoosin esiintyminen tai puuttuminen
- meioosin esiintyminen tai puuttuminen
DNA:lla on pitkä, kaksijuosteinen, spiraalimainen molekyyli, joka koostuu deoksiribonukleotideiksi kutsutuista rakennuspalikoista. Kukin deoksiribonukleotidi koostuu kolmesta osasta: 5-hiilisokerin deoksiriboosimolekyylistä, typpiemäksestä ja fosfaattiryhmästä (kuva \(\PageIndex{1}\)).
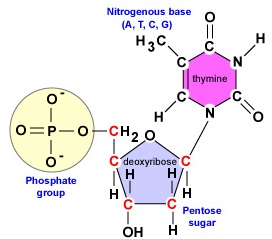
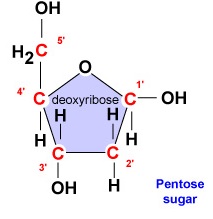
- Deoksiriboosi. Deoksiriboosi on rengasmainen 5-hiilisokeri (kuva \(\PageIndex{2}\)). Viisi hiiltä on numeroitu juoksevasti myötäpäivään sokerin ympärillä. Ensimmäiset 4 hiiltä muodostavat itse asiassa sokerin renkaan, jossa 5′ hiili irtoaa renkaan 4′ hiilestä. Nukleotidin typpiemäs on kiinnittynyt sokerin 1′ hiileen ja fosfaattiryhmä on sitoutunut 5′ hiileen. DNA-synteesin aikana entsyymi DNA-polymeraasi liittää uuden deoksiribonukleotidin fosfaattiryhmän kovalenttisesti ketjussa jo olevan nukleotidin 3′ hiileen.
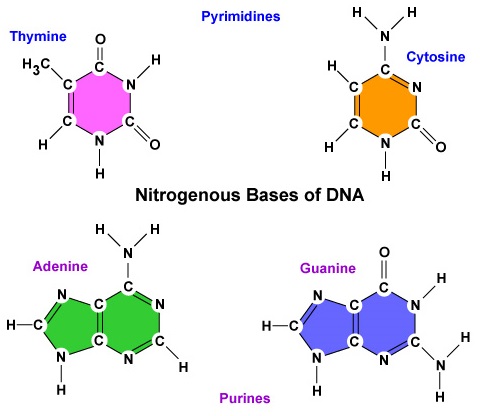
- Typpiemäs. DNA:ssa on neljä typpiemästä: adeniini, guaniini, sytosiini tai tymiini. Adeniinia ja guaniinia kutsutaan puriiniemäksiksi, kun taas sytosiinia ja tymiiniä kutsutaan pyrimidiiniemäksiksi (kuva \(\PageIndex{3}\)).
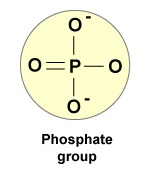
- Fosfaattiryhmä (Kuva \(\PageIndex{4}\)).
Kahden deoksiribonukleotidiketjun syntetisoimiseksi DNA:n replikaation aikana siihen osallistuvat DNA-polymeraasientsyymit pystyvät liittämään uuden nukleotidin 5′-hiilen fosfaattiryhmän vain ketjussa jo olevan nukleotidin 3′-hiilen hydroksyyliryhmään (OH-ryhmä) (kuva \(\PageIndex{2}\)). Nukleotideja yhdistävää kovalenttista sidosta kutsutaan fosfodiesterisidokseksi. Jokaisella DNA-juosteella on niin sanottu 5′-pää ja 3′-pää. Tämä tarkoittaa sitä, että kunkin DNA-juosteen toisessa päässä, jota kutsutaan 5′-pääksi , on aina fosfaattiryhmä kiinnittyneenä sen loppupään deoksiribonukleotidin 5′-hiileen (kuva \(\PageIndex{5}\)). Tämän säikeen toisessa päässä, jota kutsutaan 3′-pääksi, on aina hydroksyyli (OH) sen terminaalisen deoksiribonukleotidin 3′-hiilessä.
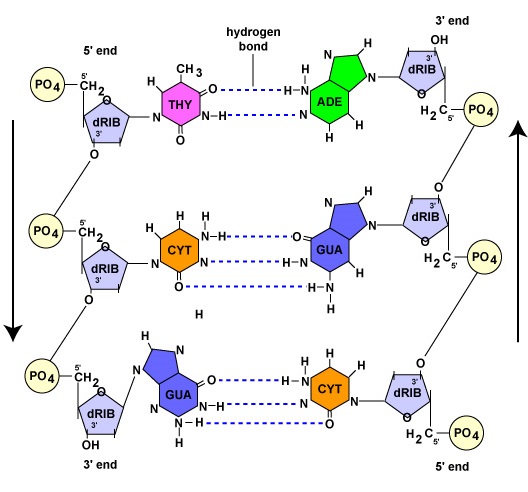
Kuten seuraavassa jaksossa nähdään, DNA:n replikaation aikana kumpikin kantasäie toimii mallina toisen säikeen synteesille komplementaarisen emäsparin avulla. Komplementaarisella emäsparinmuodostuksella tarkoitetaan DNA-nukleotideja, joiden emäs adeniini muodostaa vetysidoksia ainoastaan nukleotidien kanssa, joiden emäs on tymiini (A-T). Vastaavasti nukleotidit, joiden emäs on guaniini, voivat muodostaa vetysidoksen vain nukleotidien kanssa, joiden emäs on sytosiini (G-C). (RNA-nukleotidien tapauksessa, kuten myöhemmin nähdään, adeniininukleotidit muodostavat vetysidoksia sellaisten nukleotidien kanssa, joiden emäs on urasiili, koska tymiiniä ei esiinny RNA:ssa). Tämän sidoksen tuloksena DNA saa kierteisen muotonsa. Siksi DNA:n kahden säikeen sanotaan olevan toisiaan täydentäviä. Aina kun toisessa säikeessä on adeniinia sisältävä nukleotidi, vastakkaisessa säikeessä on tymiinin nukleotidi; aina kun toisessa säikeessä on guaniinia sisältävä nukleotidi, vastakkaisessa säikeessä on sytosiinin nukleotidi (kuva \(\PageIndex{1}\)).
Mikäli DNA:n kaksi säiettä ovat toisiaan täydentäviä, ne ovat suuntautuneet toisiinsa nähden vastakkaisiin suuntiin. Toisen säikeen sanotaan kulkevan 5′:sta 3′:een; vastakkainen DNA-säie kulkee antiparalleelisesti eli 3′:sta 5′:een (kuva \(\PageIndex{1}\)).
Vertailemme nyt lyhyesti prokaryoottisolujen genomia eukaryoottisolujen genomiin.
Prokaryoottisen (bakteerin) genomi
Bakteerin sisällä olevaa aluetta, jossa kromosomi näkyy elektronimikroskoopilla, kutsutaan nukleoidiksi. Useimpien prokaryoottien kromosomi on tyypillisesti yksi pitkä, yksittäinen molekyyli kaksisäikeistä, spiraalimaista, superkierrettyä DNA:ta, joka muodostaa fyysisen ja geneettisen kehän. Kromosomi on yleensä noin 1000 µm pitkä ja sisältää usein noin 4000 geeniä (kuva \(\PageIndex{8}\)). Escherichia coli -bakteerin, jonka pituus on 2-3 µm, kromosomi on noin 1400 µm pitkä. Jotta näin suuri makromolekyyli mahtuisi bakteerin sisälle, histonin kaltaiset proteiinit sitoutuvat DNA:han ja jakavat DNA-molekyylin noin 50 kromosomaaliseen alueeseen ja tekevät siitä tiiviimmän. DNA-topoisomeraasientsyymi, DNA-gyraasi, kierrättää sitten kromosomin tiiviiksi nipuksi, jolloin muodostuu tiivistetty, superkierretty DNA-massa, jonka halkaisija on noin 0,2 µm.
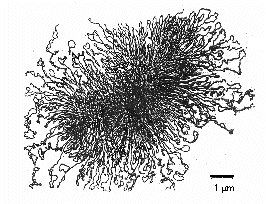
Bakteerien entsyymit, joita kutsutaan DNA-topoisomeraaseiksi, ovat välttämättömiä ympyränmuotoisen, superkierteisen bakteeri-DNA:n purkautumisessa, monistumisessa ja uudelleenkieroutumisessa (kuva \(\PageIndex{7}\)). Ne ovat myös välttämättömiä DNA:n transkriptiossa RNA:ksi, DNA:n korjauksessa ja geneettisessä rekombinaatiossa bakteereissa.
Kuva \(\PageIndex{7}\): Ympyränmuotoinen, superkierteinen prokaryoottinen DNA. Jotta suuri DNA-molekyyli mahtuisi bakteerin sisälle, DNA-topoisomeraasientsyymi nimeltä DNA-gyraasi superkierrättää kromosomin tiiviiksi nipuksi muodostaen tiivistetyn, superkierretyn DNA-massan, joka on halkaisijaltaan noin 0,2 µm.
Prokaryoottisessa nukleoidissa ei ole DNA:ta ympäröivää ydinkalvoa eikä ydinkehä eikä ydinkehä jakaannu mitoosin avulla. Sytoplasmakalvolla on merkitystä DNA:n erottumisessa bakteerien replikaation aikana. Koska bakteerit ovat haploideja (niillä on vain yksi kromosomi), niissä ei myöskään ole meioosia.
Eukaryoottinen genomi
Prokaryoottiset ja eukaryoottiset solut eroavat toisistaan hyvin paljon sekä DNA-molekyylien määrässä että organisaatiossa. Eukaryoottisoluissa on paljon enemmän DNA:ta kuin bakteereissa, ja tämä DNA on järjestäytynyt useiksi kromosomeiksi, jotka sijaitsevat tuman sisällä.
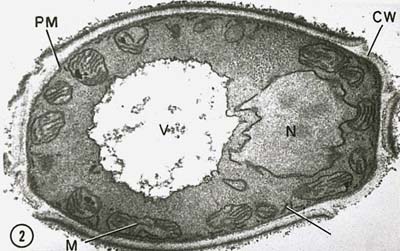
Eukaryoottisolujen tuma on ydinkalvon ympäröimä (kuva \(\PageIndex{7}\)), ja se sisältää lineaarisia kromosomeja, jotka koostuvat negatiivisesti varautuneesta DNA:sta, joka on yhdistetty positiivisesti varautuneisiin perusvalkuaisaineisiin, joita sanotaan histoneiksi, ja jotka muodostavat rakenteita, joita kutsutaan nimellä nukleosomit. Nukleosomit ovat osa niin sanottua kromatiinia, DNA:ta ja proteiineja, joista kromosomit koostuvat. Tuma jakautuu mitoosissani, ja diploidisista soluista syntyy haploidisia sukusoluja meioosin avulla.
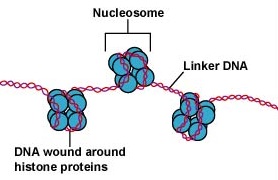
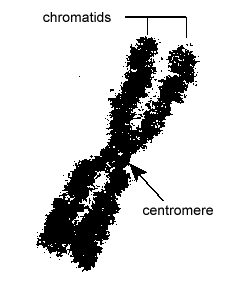
Eukaryoottisolujen DNA on pakattu hyvin organisoidusti. Se koostuu nukleosomiksi kutsutusta perusyksiköstä, joka on halkaisijaltaan 11 nm:n kokoinen helmiäismäinen rakenne, joka koostuu kahdeksan histonimolekyylin ympärille kietoutuneesta 146 emäsparista DNA:ta. Nukleosomit on yhdistetty toisiinsa noin 60 emäsparin pituisella DNA-segmentillä, jota kutsutaan linkkeri-DNA:ksi (kuva \(\PageIndex{9}\)). Toinen linkkeri-DNA:han liittyvä histoni pakkaa sitten vierekkäiset nukleotidit yhteen muodostaen halkaisijaltaan 30 nm:n nukleosomilangan. Lopuksi nämä pakatut nukleosomilangat muodostavat suuria kierteisiä silmukoita, joita histoniin kuulumattomat telineproteiinit pitävät yhdessä. Nämä telineproteiinien kierteiset silmukat muodostavat vuorovaikutuksessa kromosomeissa mitoosin aikana näkyvän tiivistetyn kromatiinin (kuva \(\PageIndex{10}\)).
Viime vuosina on havaittu, että deoksiribonukleoproteiinin rakenteellinen luonne vaikuttaa siihen, kirjoitetaanko DNA:ta RNA:ksi vai ei. Esimerkiksi kromatiinin kemialliset muutokset voivat mahdollistaa sen osien tiivistymisen tai rentoutumisen. Kun alue on tiivistynyt, geenejä ei voida transkriboida. Lisäksi kemialliset aineet voivat kiinnittyä histoniproteiineihin, joiden ympärille DNA kietoutuu, tai poistua niistä. Näiden kemiallisten ryhmien kiinnittyminen tai poistuminen histoniin määrittää, vahvistuuko tai tukahdutuuko läheisen geenin ilmentyminen.
Epigenomilla tarkoitetaan erilaisia kemiallisia yhdisteitä, jotka muokkaavat perimää tyypillisesti lisäämällä metyyli (CH3)-ryhmän nukleotidiemäkselle adeniinille tietyissä paikoissa DNA-molekyylin varrella. Tämä metylaatio voi puolestaan joko tukahduttaa tai aktivoida tiettyjen geenien transkriptiota. Periaatteessa kytkemällä geenit päälle tai pois päältä epigenomi antaa perimälle mahdollisuuden olla vuorovaikutuksessa solun ympäristön kanssa ja reagoida siihen. Epigenomi voi periytyä aivan kuten genomi.
Yhteenveto
- Deoksiribonukleiinihappo (DNA) on pitkä, kaksisäikeinen, spiraalimainen molekyyli, joka koostuu rakennuspalikoista, joita kutsutaan deoksiribonukleotideiksi.
- Deoksiribonukleotidi koostuu kolmesta osasta: 5-hiilisokerin deoksiriboosimolekyylistä, typpiemäksestä ja fosfaattiryhmästä.
- DNA:ssa on neljä typpiemästä: adeniini, guaniini, sytosiini tai tymiini. Adeniinia ja guaniinia kutsutaan puriiniemäksiksi, kun taas sytosiinia ja tymiiniä kutsutaan pyrimidiiniemäksiksi.
- Deoksiriboosi on rengasmainen 5-hiilisokeri. Viisi hiiltä on numeroitu juoksevasti myötäpäivään sokerin ympärillä. Ensimmäiset 4 hiiltä muodostavat itse asiassa sokerin renkaan, jossa 5′ hiili irtoaa renkaan 4′ hiilestä. Nukleotidin typpiemäs on kiinnittynyt sokerin 1′-hiileen ja fosfaattiryhmä on sitoutunut 5′-hiileen.
- DNA-synteesin aikana DNA-polymeraasi-entsyymi voi kiinnittää uuden deoksiribonukleotidin fosfaattiryhmän vain ketjussa jo olevan nukleotidin 3′-hiileen.
- DNA:n replikaation aikana kumpikin emäsjuoste toimii mallina toisen juosteen synteesille komplementaarisen emäsparin avulla.
- Komplementaarisella emäsparin muodostamisella tarkoitetaan sitä, että DNA:n nukleotidit, joissa on emäs adeniini, muodostavat vetysidoksia vain nukleotidien kanssa, joissa on emäs tymiini (A-T). Vastaavasti nukleotidit, joilla on emäs guaniini, voivat muodostaa vetysidoksia vain nukleotidien kanssa, joilla on emäs sytosiini (G-C).
- Mikäli DNA:n kaksi säiettä ovat komplementaarisia, ne ovat suuntautuneet vastakkaisiin suuntiin toisiinsa nähden. Toisen säikeen sanotaan kulkevan 5′ – 3′; DNA:n vastakkainen säie kulkee antiparalleelisesti eli 3′ – 5′.
- Prokaryoottisoluissa DNA:ta ei ympäröi ydinkalvo. Prokaryoottisoluista puuttuvat mitoosi ja meioosi.
- Jotta näin suuri makromolekyyli mahtuisi bakteerin sisälle, histonin kaltaiset proteiinit sitoutuvat DNA:han, jolloin DNA-molekyyli jakautuu noin 50 kromosomialueeseen ja siitä tulee tiiviimpi. Tämän jälkeen DNA-gyraasi-niminen entsyymi kierrättää jokaisen domainin itsensä ympärille muodostaen tiivistetyn, superkierretty DNA-massan. DNA-gyraasi-niminen topoisomeraasi katalysoi bakteereissa esiintyvän ympyränmuotoisen DNA:n negatiivista superkäämitystä. Topoisomeraasi IV puolestaan osallistuu superkierteisen ympyränmuotoisen DNA:n rentouttamiseen, mikä mahdollistaa toisiinsa kytkeytyneiden tytärkromosomien erottamisen bakteerien DNA:n replikaation lopussa.
- Eukaryoottisolujen DNA on pakattu perusyksiköihin, joita kutsutaan nukleosomeiksi, helmiä muistuttaviksi rakenteiksi, jotka koostuvat DNA:sta, joka on kietoutunut kahdeksan histonimolekyylin ympärille. DNA on järjestäytynyt useiksi kromosomeiksi, jotka sijaitsevat ytimessä, jota ympäröi ydinkalvo. Tuma jakautuu mitoosin avulla ja sukusolut syntyvät meioosin avulla sukupuolisesti lisääntyvillä eukaryooteilla.
- Deoksiribonukleoproteiinin rakenteellinen luonne vaikuttaa siihen, transkriboituvatko DNA:t RNA:ksi vai eivät. Esimerkiksi kromatiinin kemialliset muutokset voivat mahdollistaa sen osien tiivistymisen tai rentoutumisen. Kun alue on tiivistynyt, geenejä ei voida transkriboida. Lisäksi kemialliset aineet voivat kiinnittyä histoniproteiineihin, joiden ympärille DNA kietoutuu, tai poistua niistä. Näiden kemiallisten ryhmien kiinnittyminen tai poistuminen histoniin määrää sen, onko läheisen geenin ilmentyminen voimistunut vai tukahdutettu.
Tekijät ja attribuutit
-
Dr. Gary Kaiser (COMMUNITY COLLEGE OF BALTIMORE COUNTY, CATONSVILLE CAMPUS)