En mikrobiologisk biorealmside om slægten Rhodospirillum rubrum
Klassifikation
Taxa af højere orden
Kongedømme: Bakterier
Familie: Proteobakterier
Klasse: Proteobacteria
Klasse: Alphaproteobacteria
Order: Proteobakterier
Ord: Rhodospirillales
Familie: Rhodospirillales
Familie: Rhodospirillaceae
Genus: Rhodospirillaceae
Genus: Rhodospirillum
Species
Rhodospirillum rubrum
Beskrivelse og betydning
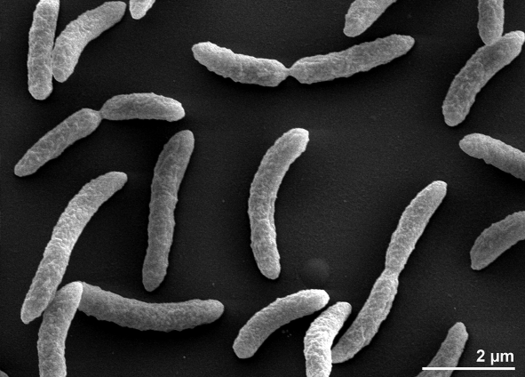
Rhodospirillum rubrum er en gramnegativ, mesofil proteobakterie. Dens optimale væksttemperatur er 25-30 grader Celsius. Den har flerlagede ydre hylstre, som for det meste indeholder umættede, men enkelte mættede fedtstoffer i dens cellevæg. R. rubrum er en spirilla, hvilket betyder, at den har en spiralformet form. Den er polært flagelleret og er derfor motil. Dens længde er 3-10 um, med en bredde på 0,8-1,0 um.
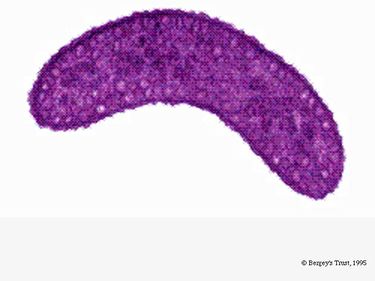
R. rubrum er en fakultativ anaerobe. Afhængigt af tilstedeværelsen af ilt kan den undergå alkoholisk gæring eller aerob respiration. Den er også i stand til fotosyntese og indeholder carotenoid og baterioklorofyl i sine chromatophorpartikler. Disse molekyler hjælper med at absorbere lys og omdanne det til energi og giver den også sin tydelige purpurrøde farve under anaerobe forhold. R. rubrum er farveløs under aerobe forhold.
Og selv om fotosyntesen er aktiv under aerobe forhold, er den generelt undertrykt i nærvær af O2. Svovl er et vigtigt biprodukt af fotosyntesen, ikke O2. R. rubrum kan vokse heterotrofisk eller autotrofisk, når den er fotosyntetisk. I modsætning til mange planter indeholder R. rubrum ikke klorofyl a (absorptionsspektret 430-662 nm). Den indeholder dog klorofyl b (absorptionsspektre 660-680 nm) og bakterioklorofyls (800-925 nm). Dette gør det muligt for den at udnytte mere energi fra de elektromagnetiske spektrer. R. rubrum oxiderer også carbonmonoxid (CO) med brintgas som slutprodukt af vejen og kan bruge sulfid i lave koncentrationer som elektrondonor i kuldioxidreduktion. Derudover er den en kvælstoffikserende bakterie; den bruger nitrogenase til at omdanne atmosfærisk kvælstofgas til ammoniak (Munk et al, 2011).
Der er flere anvendelser af R. rubrum inden for bioteknologi. Den er et modelsystem for omdannelse af lys til kemisk energi og for dens kvælstoffikserende veje. Den er også genstand for undersøgelser af strålingsresistens. Den kan også anvendes på flere måder til forbrug. Proteobakterierne kan være en kilde til dyrefoder og landbrugsgødning. En anden vigtig rolle i forskningen omfatter produktion af vitaminer. Der forskes også i produktion af biologisk plastik fra forløbere af polyhydroxy-buttersyre. R. rubrum kan også være en bidragyder i biologiske brintbrændstoffer, hovedsagelig gennem dens udvikling af enzymet nitrogenase.
Genomstruktur
Genomets sekventering er afsluttet. Det består af et cirkulært kromosom med 4 352 825 basepar, hvoraf 65 % er guanin-cystosinpar. Der er også et plasmid med 53.732 basepar med 60% guanin-cytosinpar. I alt er der 3.850 proteinkodende gener og 83 RNA-gener. Af disse gener er 6,9 % transkriptionsrelaterede; 4,6 % translation, ribosomstruktur og biosyntese; 4,0 % replikation, rekombination og reparation; 7,9 % signaltransduktionsmekanismer; 5,9 % cellevæg- og membranbiogenese; 6,6 % energiproduktion og -omdannelse; 5,0 % kulhydrattransport og -metabolisme; 9,5 % kulhydrattransport og -metabolisme.9 % transport og metabolisme af aminosyrer; 4,7 % transport og metabolisme af coenzymer; 3,7 % transport og metabolisme af lipider; og 6,5 % transport og metabolisme af uorganiske ioner(Munk et al, 2011).
Ekologi
Da Rhodospirillum rubrum kan vokse både aerobt og anaerobt, er den i stand til at leve under en lang række forskellige forhold. R. rubrum findes i mange naturlige akvatiske miljøer som f.eks. damme, søer, vandløb og stående vand (Reslewic et. al, 2005). R. rubrum findes også ofte i mudder og spildevand (Brock et al., 2000). Undersøgelser har vist, at R. rubrum kan foretage store ændringer i sin kemiske sammensætning for at tilpasse sig til forskellige miljøer (Cohen-Bazire og Kunisawa, 1963). R. rubrum foretrækker at vokse i levesteder med en pH-værdi på 6,8-7,2 (Bergey og Holt, 1994). Undersøgelser har også vist, at R. rubrum har en optimal væksttemperatur på 22-35 grader Celcius (Weaver, 1971).
Cellestruktur og stofskifte
R. rubrum er en alsidig organisme, der kan skaffe sig energi gennem en række forskellige mekanismer. Respiration og fotosyntetiske mekanismer eksisterer sammen, og deres aktivitet er afhængig af tilstedeværelsen af lys og energi. R. rubrum kan vokse i mørke kemotropiske miljøer med tilstedeværelse af ilt eller kan vokse i et fototropisk miljø uden ilt.
Fotosyntesen i R. rubrum begynder med donation af et hydrogen fra et organisk substrat til et oxiderende substrat. Bakterioklorofyl og cartenoider, de fotoreaktive pigmenter, der findes i cellemembranen, er bundet til chromatophorer (Schachman, Pardee og Stanier, 1952). Chromatophorerne er fladtrykte skiver, der indeholder cholinphospholipider, cardio lipin og galactosyl diglycerider (Benson, Wintermans og Wiser, 1959; Benson 1961). Desuden indeholder chromatophorerne en komplet elektrontransportkæde, der omfatter forskellige cytokromer, flavin og pyridinnukleotider (Newton og Newton, 1957; Hulcher og Conti, 1960; Kamen, 1961). Cellemembranen indeholder også maskineri til ATP-syntese, herunder en ATP-synthetase.
I tilstedeværelse af ilt er R. rubrum i stand til aerobt at respirere ved hjælp af en traditionel elektrontransportkæde med NAD+/NADH som den primære elektronbærer (Keister og Minton, 1969). Blandt elektrontransportproteinerne er cytochrom C428 (Chance og Smith, 1955). Aerob respiration hæmmes imidlertid af fotosyntese (Oelze og Weaver, 1971).
R. rubrum er også i stand til anaerob respiration. Dens ikke-oxygeniske terminale elektronacceptorer kan omfatte dimethylsulfoxid og trimethylaminoxid. Tilstedeværelsen af disse elektronacceptorer gør det muligt for substrater som f.eks. succinat, malat og acetat at understøtte væksten af R. rubrum. Disse acceptorer er dog kun ca. 33-41 % så effektive som ilt med hensyn til energibesparelse (Schultz og Weaver, 1982).
Dertil kommer, at R. rubrum har den unikke evne til at oxidere carbonmonoxid ved hjælp af carbonmonoxiddehydrogenase. Denne oxidationsvej ender med reduktion og brint og produktion af brintgas.
Selv uden en terminal elektronacceptor gennemfører R. rubrum blandet syrefermentering. De vigtigste produkter fra pyruvatfermentering er acetat, formiat, kuldioxid og hydrogen. I tilstedeværelse af bicarbonat-ioner kan fructose fermenteres. Slutprodukterne af denne vej er de samme som for pyruvat, men med tilføjelse af succinat og propionat (Schultz og Weaver 1982).
Ud over at have evnen til at fiksere kuldioxid (Schon og Biedermann, 1972) er en identificerende anabolsk egenskab hos R. rubrum dens evne til at fiksere kvælstof. Under mørke forhold med tilstedeværelse af fructose bruger R. rubrum nitrogenase til at fiksere kvælstofgas til ammonium. Den indeholder både Fe-Mo og Fe-only nitrogenaser.
Patologi
R. rubrum har ikke vist sig at inficere mennesker eller dyr.
Aktuel forskning
Stråleresistens
Rhodospirillum rubrum har for nylig givet indsigt i stråleresistens. Felice Mastroleo et al. har i Insight into the radiotolerance of the life support bacterium Rhodospirillum rubrum S1H by means of phenotypic and transcriptomic methods (2010) karakteriseret R. rubrums strålingstolerance.
Ved at udsætte bakterien for subdødelige doser af Cobalt-60 gammastråler, en type ioniserende stråling, blev R. rubrum-markørgener, der er specifikke for ioniserende stråling, karakteriseret i organismen. Disse gener steg i ekspression, efterhånden som “restitutionstiden” forløb efter en given gammastrålebehandling. Analysen af genekspressionen i R. rubrum efter gammastråling skaber mulighed for yderligere mikrobiologisk forskning, som kan føre til fremskridt inden for anvendt strålingsresistens i fremtiden.
Biologisk nedbrydelige plastmaterialer
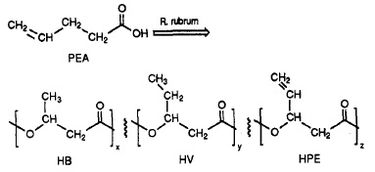
R. rubrum har vist sig at kunne producere en klasse af bionedbrydelige plastmaterialer, polybeta-hydroxyalkanoater (PHA’er). Anvendelserne af denne forskning kan løse mange problemer, der er forårsaget af syntetisk, ikke-nedbrydelig plast, som har haft en negativ indvirkning på miljøet. Et forskningseksperiment udført af Herbert. W. Ulmer et al. har beskrevet produktionen af PHA’er indeholdende 3HV-enheder ved hjælp af Rhodospriillum rubrum (1993). Ulmer anvendte 4-pentenoic acid (PEA) som vækstmedium for R. rubrum. Når bakterien blev anbragt under “PHA-producerende forhold”, blev der syntetiseret PHA’er, der indeholdt 3 gentagne underenheder (bemærk: R = substituentgruppe): HB (R = methyl), HV (R = ethyl), HPE (R = vinyl) (se figur 1).
Syntesen af funktionelle, bionedbrydelige plaststoffer ved hjælp af bakterier har potentiale til at reducere den globale forurening og brændstofudgifterne fra plastfabrikker. R. rubrum kan bruges som et mikrobiologisk værktøj til at fremstille miljøvenlige biopolymerer.
Renoverbar energi
Den stigende bekymring over miljøforurening fra afbrænding af fossile brændstoffer har ført til en stigning i forskningen af Rhodospirillum rubrum. Ved at anvende R. rubrum som kilde til brint til brug i vedvarende brintbrændstoffer løses flere problemer, der er skabt af forbruget af fossile brændstoffer. Brint oxideres til vand som et forbrændingsprodukt i modsætning til fossile brændstoffer, der skaber forurenende stoffer som CO, CO2, SOx og NOx. R. rubrum kan producere brint ved at udnytte følgende “vand-gasforskydningsreaktion” (Najafpour et al., 2004):
CO + H2O → H2 + CO2
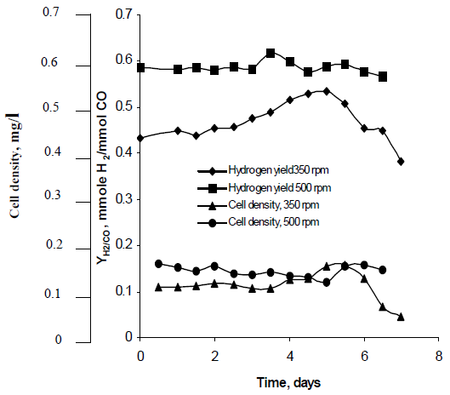
Najafpours forskningsmetode observerede, hvordan omrøring, der blev skabt af sæt turbinehjul indstillet med hastigheder på 350 rpm samt 500 rpm, påvirkede R. rubrums brintproduktionshastighed. Hans forskning viste, at brintbrændstof har potentiale til at dæmme op for den globale opvarmning og give en miljøvenlig teknologisk løsning på brændstofproduktion og -forbrug. Hans undersøgelse viste, at “højere omrøringshastighed gav højere brint samt celletæthed” af R. rubrum (se figur 2).
I en anden mikrobiologisk undersøgelse, udført af Matthew Ross Melnicki, blev det fundet, at R. rubrum giver potentiale for vedvarende energi, såsom brintbrændstof, via brintsyntese. Dens biologiske veje kan producere brintniveauer med høj hastighed, når den samkultiveres med Chlamydomonas reinhardtii, en algeart, som R. rubrum kan vokse harmonisk sammen med. Melnicki foreslog en ukonventionel tilgang til samkultivering til syntese af brint, som producerede forbedrede niveauer af brintsyntese, som ikke kan opnås, hvis samkultiveringen er afkoblet. C. reinhardtii var begunstiget ved højere lysintensiteter, mens R. ruburm var mere produktiv under lavere lysintensiteter, hvilket fremhæver fordelene ved deres samkultivering.
Ved anvendelse af en fed-batch teknik til dyrkning af R. rubrum blev det fundet, at inokulerede kulturer producerede brint under dens eksponentielle vækstfase, og syntesen af brint fortsatte med at blomstre kontinuerligt i 70 timer. Da væksten ophørte, tilføjede Melnicki succinat, der fungerer som en elektrondonor i den metaboliske vej, og brintudviklingen blev genoptaget, selv om bakterierne nu ikke længere voksede. Det er dog vigtigt at bemærke, at der ved kontinuerlig tilsætning af succinat blev fundet reducerede udviklingshastigheder af brint. Når der blev tilsat yderligere vækstmedier, steg brintproduktionshastighederne igen.
Melnicki udtalte også, at “selv om vækst ikke er nødvendig for brintproduktion, fastslår dette arbejde nødvendigheden af cellevækst for at opretholde maksimale hastigheder, hvilket tyder på den industrielle egnethed af en semi-kontinuerlig kulturstrategi” (Melnicki, 2009).
Cool Factor
Genomsekvensen af Rhodospirillum rubrum er kun den anden af Rhodospirillum-slægten, der er blevet offentliggjort, efter R. centenum (Munk et al, 2011).

Et interessant aspekt ved Rhodospirillum rubrum er dens evne til at skifte farve under forskellige forhold. Den er en anoxygen fototrof, men producerer ikke ekstracellulær ilt. I stedet producerer den elementært svovl under lyshøstning. Dette skyldes, at R. rubrum mangler lyshøstningskompleks 2 (Munk et al., 2011). Bakterien anvender bakterioklorofyl, hvilket gør den i stand til at absorbere bølgelængder af lys, som planter ikke udnytter. Bakterioklorofyl absorberer bølgelængder på 800-925 nm.
Under aerobe forhold er den fotosyntetiske aktivitet hos R. rubrum genetisk undertrykt, og bakterien bliver farveløs. Under anaerobe forhold aktiveres fotosyntesen imidlertid, og bakterien får en tydelig purpurrød farve (se billede 5). Dette skyldes aktivitet i chromatophorerne, som findes i membranen (Munk et al., 2011). Chromatophorerne indeholder bakteriens bakterioklorofylpigmenter og undergår absorptionsændringer under aktivering af fotosyntesen.
Benson, A. “Chloroplast lipid metabolism.” I Symposium nr. VI. 5. internationale kongres om biokemi. 1961. Oxford: Pergamon Press. (I trykken.)
Benson, A., Wintermans, J. og Wiser, R. “Chloroplast lipids as carbohydrate reservoirs.” Plant Physiology. 1959. Volume 34. p. 815.
Bergey, D. og Holt, J. Bergey’s manual of determinative bacteriology. Niende udgave. Baltimore. Lippincott Williams and Wilkins. 1994. s. 359.
Brock, T.D., M.T. Madigan, J.M. Martinko, og J Parker. Biologi af mikroorganismer. 9. udgave. Upper Saddle River: Prentice Hall, 2000. s. 459.
Chance, B. og Smith, L. “Respiratoriske pigmenter i Rhodospirillum rubrum.” Nature. Bind 174. s. 803-806.
Cohen-Bazire, G. og Kunisawa, R. “The Fine Structure of Rhodospirillum Rubrum .” Journal of Cell Biology. 1963. Volume 16. p. 401-419.
Hulcher, M. og Conti, S. “Cytochronies in chlorophyll-containing particles of Chromatium and Chlorobium thiosuphatophilum.” Biochemistry and Biophysics Research Communications. 1960. Volume 3. p. 497.
Kamen, M. “The Haem Proteins of Photosynthetic Tissues.” I Symposium nr. VI. 5. internationale kongres om biokemi. 1961. Oxford: pergamon Press. (I trykken.)
Keister, D. og Minton, N. “Energy-linked reactions in photosynthetic bacteria. 3. Yderligere undersøgelser af energibundet nikotinamid-adenin-dinukleotidreduktion ved Rhodospirillum rubrum chromatophorer.” Biochemistry. 1969. Volume 8. p. 167-173.
Mastroleo, F., Monsieurs, P., and Leys, N. “Insight into the radiotolerance of the life support bacterium Rhodospirillum rubrum S1H by means of phenotypic and transcriptomic methods”. COSPAR’s videnskabelige forsamling. 2010. s. 4.
Melnicki, M. R. “Modes of hydrogen production in the photosynthetic bacterium, Rhodosphirillum rubrum”. University of California, Berkeley. 2009.
Munk, C., Copeland, A., Lucas, C., Lapidus, A., et al. “Complete genome sequence of Rhodospirillum rubrum type strain (S1T)”. Standards in Genomic Sciences. 2011. Volume 4. pp. 293-302.
Najafpour, G., Ismail, K., Younesi, H., Mohamed, A. R., and Kamaruddin, A. H. “Hydrogen as clean fuel via continuous fermentation by anaerobic photosynthetic bacteria, Rhodospirillum rubrum”. African Journal of Biotechnology. 2004. Volume 3. pp. 503-507.
Newton, J. og Newton, G. “Composition of the photoactive subcellular particles from Chromatium.” The Archives of Biochemistry and Biophysics. 1957. Volume 71. p. 250.
Oelze, J. og Weaver, P. “The adjustment of photosynthetically grown cells of Rhodospirillum rubrum to aerobic light conditions.” The Archives of Microbiology. 1982. Volume 79. p. 108-121.
Reslewic, S., Shigou, Z., Place, M., Zhang, Y., Briska A., Goldstein, S., Churas, C., Runnheim, R., Forrest, D., Lim, A., Lapidus, A., Han, C., Roberts, G., og Schwartz, D. “Whole-Genome Shotgun Optical Mapping of Rhodospirillum rubrum .” Anvendt og miljømæssig mikrobiologi. 2005. Volume 71. s. 5511-5522.
Schachman, H., Pardee, A., og Stanier, R. “Studies on the macromolecular organization of microbial cells.” The Archives of Biochemistry and Biophysics. 1952. Volume 38. p. 245-260.
Schön, G. og Biedermann, M. “Synthese af flygtige syrer ved fermentering af pyruvat og fructose i anaerobe mørke kulturer af Rhodospirillum rubrum.” Archiv fur Mikrobiologie.1972. Volume 85. p. 77-90.
Schultz, J. and Weaver, P. “Fermentation and anaerobic respiration by Rhodospirillum rubrum and Rhodopseudomonas capsulate.” The Journal of Bacteriology. 1982. Volume 149. p. 181-190.
Ulmer, H., Gross, R., Posada, M., Weisbach, P., Fuller, R. C., and Lenz, R. “Bacterial production of poly(.beta.-hydroxyalkanoates) containing unsaturated repeating units by Rhodospirillum rubrum”. Makromolekyler. 1994. Volume 27. p. 1675-1679.
Weaver, P. “Temperature-Sensitive Mutations of the Photosynthetic Apparatus of Rhodospirillum rubrum”. Proceedings of the National Academy of Sciences of the United States of America. 1971. Volume 68. p. 136-138.
Redigeret af studerende af Dr. Iris Keren: Allison Almeida, Matthew Cozzolino, Trung Nguyen og Jessica Wong.
NEUF2011