
Plantcelle
En eukaryote celles kerne styrer direkte eller indirekte stort set alle cellulære fysiologiske aktiviteter, herunder initiering, regulering og afslutning af enzymatiske hændelser.
Den er også opbevaringssted for genetisk information (genomet), idet den huser og beskytter kromosomerne og de gener, som de bærer. I alle eukaryote celler er kernen generelt den største og mest centralt placerede cellestruktur, selv om visse former for vacuoler i planteopbevaringsceller kan være større og mere iøjnefaldende.
Kernen, der kommer af det græske ord nucle (der betyder “grube” eller “kerne”), er cellens kommando- og kontrolcenter. De seks grundlæggende funktioner i kernen er for det første at beskytte og opbevare generne og i sidste ende beskytte den desoxyribonukleinsyre (DNA), som generne er organiseret på, mod resten af cellen.


For det andet at organisere generne i kromosomer for at lette deres bevægelse og fordeling under celledeling; for det tredje at organisere afviklingen af DNA’et under kopieringen af generne til produktion af tusindvis af proteiner.
Fjerde, at fremstille og transportere regulerende molekyler, for det meste enzymer og andre genprodukter, til cytoplasmaet; femte, at fremstille underenheder af ribosomer; og sjette, at reagere på hormoner og andre kemiske signaler, der modtages via kerneporerne.
Komponenter
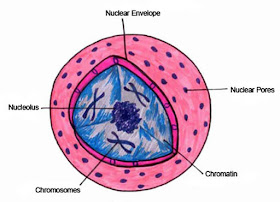
Strukturelt består kernen af flere forskellige dele: en kernehinde, kerneplasma, kromatin og en eller flere underorganeller kaldet nukleoler. Kerneomslaget danner en beskyttende barriere, der isolerer kernen fra cellens cytoplasma
Kernehyllen består af to enhedsmembraner (en dobbelt-enhedsmembran), som strukturelt ligner cellens andre membraner. Den ydre membran er tæt forbundet med cellens endoplasmatiske recticulum (ER) og kan være sammenhængende med det. Ligesom cytoplasmaets ru ER har den ydre kernemembran ribosomer indlejret i den.
Kernediagram
Nogle forskere antyder faktisk, at kerneomslaget blot er en lokaliseret og specialiseret udgave af ER’en. Den indre kernemembran er beklædt med et fibrøst lag, kaldet kernelamina, som giver styrke og struktur til kerneformen og kan også fungere som bindingssted for noget kromatin.
Med mellemrum er kerneomslaget perforeret af små porer, der fungerer som kommunikationskanaler til kontrolleret udveksling af materialer mellem kernen og cytoplasmaet. Samlet set dækker kerneporerne omkring 10 procent af kerneoverfladen.
Hver kerneporer er et kompleks bestående af en central pore, der er blevet anslået til at være 30-100 nanometer i diameter. De selektivt permeable kerneporer fungerer som ind- og udgangsveje for en række vandopløselige molekyler, for det meste kerneprodukter, såsom ribosomunderenheder, messenger RNA (ribonukleinsyre)-molekyler og kromosomale proteiner.
Protoplasmaet inden for kernen kaldes nukleoplasma. Ligesom cytoplasmaet består det af en geléagtig blanding af stoffer og organeller, men det adskiller sig ved at have en højere koncentration af nukleotider og andre organiske molekyler, der bruges i syntesen af DNA og RNA.
De vigtigste strukturer inden for nukleoplasmaet omfatter DNA’et og normalt én organel – men nogle gange flere – kaldet nukleolus. Bortset fra under celledeling forekommer DNA-molekylerne som et netværk af udrullede fibre kaldet kromatin. Under celledelingen vikler og supervikler molekylære DNA-strenge sig om histonproteiner for at kondensere sig og danne kromosomer.
At antallet af kromosomer, der findes i kernen, er specifikt for hver enkelt plante- og dyreart. Mennesker har f.eks. 46 kromosomer, tobak har 48 kromosomer, majs har 20 kromosomer, gulerødder har 18 kromosomer, og ærter har 14 kromosomer.
Nukleolus er den største synlige organel i kernen. Den er typisk forbundet med specifikke regioner af kromosomer, kaldet nukleære organizerregioner, som indeholder gener, der styrer syntesen af ribosomale underenheder.
De vigtigste produkter af nukleolusaktivitet er enhederne af ribosomalt RNA (rRNA). Disse underenheder kompleksiseres til sidst med ribosomale proteiner og transporteres fra kernen til cytoplasmaet af særlige transportproteiner.
Andre steder i kernen, kaldet funktionelle domæner, styrer syntesen af budbringer- (pre-mRNA), transfer- (tRNA) molekyler. Når disse molekyler er dannet, dannes de derefter komplekser med proteiner og transporteres som nukleoproteiner til cytoplasmaet.
Celledeling
Selv om kernen tilsyneladende er både stabil og holdbar, forsvinder den fra det normale syn og omdannes under celledeling hos næsten alle planter undtagen gær, som beholder en klart defineret kerne under hele delingsprocessen.
I andre eukaryote planteceller forsvinder kernen tidligt under mitosens profase, hvor kernehulen enzymatisk fragmenteres i små, næsten usynlige vesikler. Disse samles ikke igen før de sidste hændelser i telofasen, hvor de dannes på ny omkring kromosomerne og kontrolleres af dattercellens lamina.