Nukleinsyrernes nitrogenholdige baser, som er vigtige bestanddele af nukleotider, er organiske molekyler og er navngivet sådan, fordi de indeholder kulstof og nitrogen. De er baser, fordi de indeholder en aminogruppe, der har potentiale til at binde et ekstra hydrogen og dermed nedsætter hydrogenionkoncentrationen i omgivelserne, hvilket gør dem mere basiske. Hver nukleotid i DNA indeholder en af fire mulige nitrogenbaser: adenin (A), guanin (G), cytosin (C) og thymin (T). RNA-nukleotider indeholder også en af fire mulige baser: adenin, guanin, cytosin og uracil (U) i stedet for thymin.
Adenin og guanin er klassificeret som puriner. Den primære struktur af et purin er to kulstof-stikstofringe. Cytosin, thymin og uracil er klassificeret som pyrimidiner, som har en enkelt kulstof-kvælstofring som primær struktur (figur 1). Hver af disse grundlæggende kulstof-stivstofringe har forskellige funktionelle grupper knyttet til sig. I molekylærbiologisk forkortelse er de nitrogenholdige baser ganske enkelt kendt under symbolerne A, T, G, C og U. DNA indeholder A, T, G og C, mens RNA indeholder A, U, G og C.
Den pentose sukker i DNA er deoxyribose, og i RNA er sukkerstoffet ribose (figur 1). Forskellen mellem sukkerstofferne er tilstedeværelsen af en hydroxylgruppe på det andet kulstof i ribose og hydrogen på det andet kulstof i deoxyribose. Kulstofatomerne i sukkermolekylet er nummereret som 1′, 2′, 3′, 4′ og 5′ (1′ betyder “et primtal”). Phosphatresten er knyttet til hydroxylgruppen på 5′-kulstoffet i det ene sukkerstof og hydroxylgruppen på 3′-kulstoffet i sukkeret i det næste nukleotid, hvorved der dannes en 5′-3′-fosfodiesterbinding. Phosphodiesterbindingen dannes ikke ved en simpel dehydreringsreaktion som de andre bindinger, der forbinder monomerer i makromolekyler: dens dannelse indebærer fjernelse af to fosfatgrupper. Et polynukleotid kan have tusindvis af sådanne fosfodiesterbindinger.
DNA Double-Helix Structure
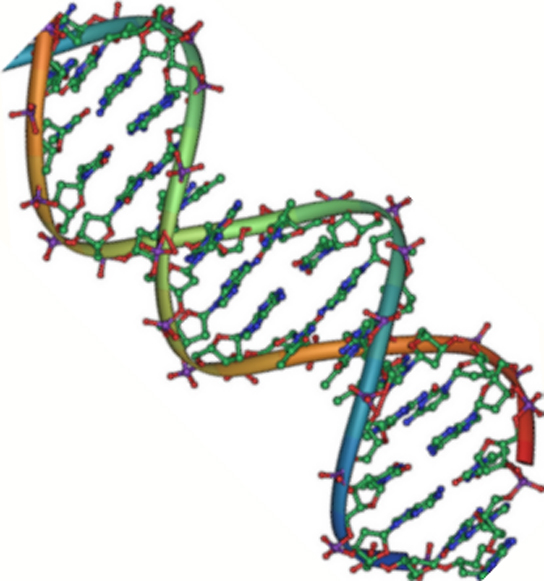
Figur 2. DNA er en antiparallel dobbelthelix. Phosphatryggen (de krumme linjer) er på ydersiden, og baserne er på indersiden. Hver base interagerer med en base fra den modsatte streng. (credit: Jerome Walker/Dennis Myts)
DNA har en dobbelt-helix-struktur (figur 2). Sukkeret og fosfatet ligger på ydersiden af helixen og danner DNA’s rygsøjle. De nitrogenholdige baser er stablet i det indre, som trinene på en trappe, i par; parrene er bundet til hinanden ved hjælp af hydrogenbindinger. Hvert basepar i dobbelthelixen er adskilt fra det næste basepar med 0,34 nm.
De to strenge i helixen løber i modsatte retninger, hvilket betyder, at 5′-kulstofenden af den ene streng vil vende mod 3′-kulstofenden af den tilsvarende streng. (Dette kaldes antiparallel orientering og er vigtigt for DNA-replikation og i mange nukleinsyreinteraktioner.)
Kun visse typer baseparring er tilladt. F.eks. kan et bestemt purin kun parre sig med et bestemt pyrimidin. Det betyder, at A kan parre sig med T, og at G kan parre sig med C, som vist i figur 3. Dette er kendt som reglen om basekomplementaritet. Med andre ord er DNA-strengene komplementære til hinanden. Hvis sekvensen på den ene streng er AATTGGCC, vil den komplementære streng have sekvensen TTAACCGG. Under DNA-replikationen kopieres hver streng, hvilket resulterer i en datter-DNA-dobbeltspiral, der indeholder en forældre-DNA-streng og en nyligt syntetiseret streng.
Praktikspørgsmål
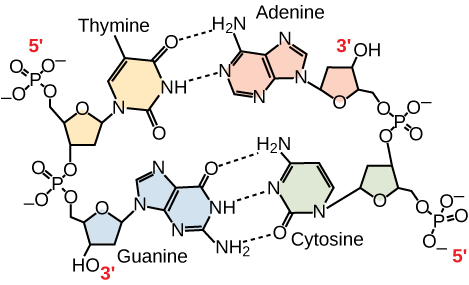
Figur 3. I et dobbeltstrenget DNA-molekyle løber de to strenge antiparallelt til hinanden, således at den ene streng løber 5′ til 3′ og den anden 3′ til 5′. Phosphatryggen er placeret på ydersiden, og baserne er i midten. Adenin danner hydrogenbindinger (eller basepar) med thymin, og guanin danner basepar med cytosin.
Der sker en mutation, og cytosin erstattes med adenin. Hvilken indvirkning tror du, at dette vil have på DNA-strukturen?
RNA
Ribonukleinsyre, eller RNA, er hovedsageligt involveret i processen med proteinsyntese under ledelse af DNA. RNA er normalt enkeltstrenget og består af ribonukleotider, der er forbundet af phosphodiesterbindinger. Et ribonukleotid i RNA-kæden indeholder ribose (pentosesukker), en af de fire nitrogenbaser (A, U, G og C) og fosfatgruppen.
Der findes fire hovedtyper af RNA: messenger RNA (mRNA), ribosomalt RNA (rRNA), transfer RNA (tRNA) og microRNA (miRNA). Det første, mRNA, bærer budskabet fra DNA, som styrer alle celleaktiviteterne i en celle. Hvis en celle har brug for, at et bestemt protein skal syntetiseres, “tændes” genet for dette produkt, og budbringer-RNA’et syntetiseres i kernen. RNA-basesekvensen er komplementær til kodningssekvensen i det DNA, som den er blevet kopieret fra. I RNA er basen T imidlertid ikke til stede, og i stedet er basen U til stede. Hvis DNA-strengen har sekvensen AATTGCGC, er sekvensen for det komplementære RNA UUAACGCG. I cytoplasmaet interagerer mRNA’et med ribosomer og andre cellulære maskiner (figur 4).
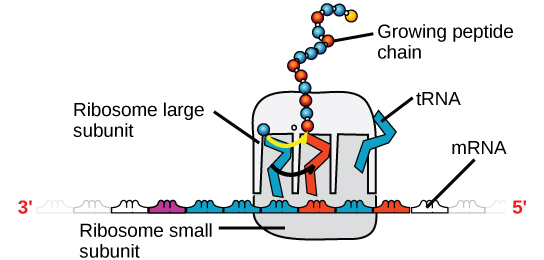
Figur 4. Et ribosom består af to dele: en stor underenhed og en lille underenhed. Det mRNA sidder mellem de to underenheder. Et tRNA-molekyle genkender et kodon på mRNA’et, binder sig til det ved komplementær baseparring og tilføjer den korrekte aminosyre til den voksende peptidkæde.
Det mRNA læses i sæt af tre baser, der kaldes kodoner. Hvert codon koder for en enkelt aminosyre. På denne måde læses mRNA’et, og proteinproduktet fremstilles. Ribosomalt RNA (rRNA) er en vigtig bestanddel af ribosomerne, som mRNA’et bindes på. RRNA sikrer den korrekte tilpasning af mRNA’et og ribosomerne; ribosomernes rRNA har også en enzymatisk aktivitet (peptidyltransferase) og katalyserer dannelsen af peptidbindinger mellem to tilpassede aminosyrer. Transfer-RNA (tRNA) er en af de mindste af de fire RNA-typer og er normalt 70-90 nukleotider lang. Det transporterer den korrekte aminosyre til det sted, hvor proteinsyntesen finder sted. Det er baseparringen mellem tRNA’et og mRNA’et, der gør det muligt at indsætte den korrekte aminosyre i polypeptidkæden. microRNA’er er de mindste RNA-molekyler, og deres rolle omfatter regulering af genekspressionen ved at gribe ind i ekspressionen af visse mRNA-meddelelser.