En sida om mikrobiella biorealmer om släktet Rhodospirillum rubrum
Klassificering
Taxa av högre ordning
Kungarike: Bakterier
Familj: Proteobakterier
Klass: Alphaproteobacteria
Ord: Rhodospirillales
Familj: Rhodospirillaceae
Genus: Rhodospirillum
Species
Rhodospirillum rubrum
Beskrivning och betydelse
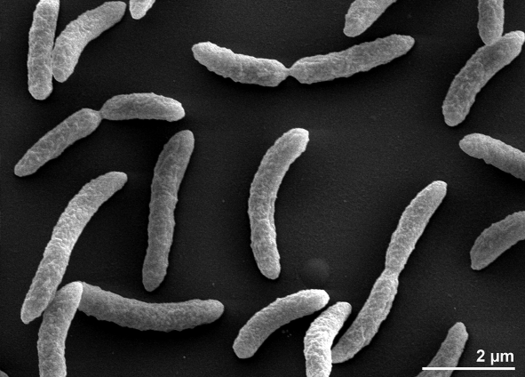
Rhodospirillum rubrum är en gramnegativ, mesofil proteobakterie. Dess optimala tillväxttemperatur är 25-30 grader Celsius. Den har flerskiktade yttre höljen som innehåller mestadels omättade, men en del mättade fetter i sin cellvägg. R. rubrum är en spirilla, vilket innebär att den har en spiralform. Den är polärt flagellerad och därför rörlig. Dess längd är 3-10 um, med en bredd på 0,8-1,0 um.
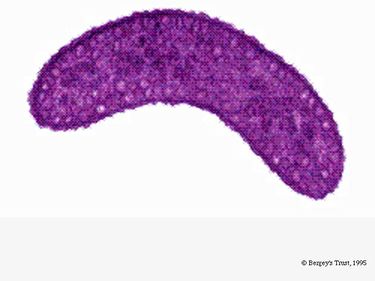
R. rubrum är en fakultativ anaerob. Beroende på närvaron av syre kan den genomgå alkoholjäsning eller aerob andning. Den har också förmåga till fotosyntes och innehåller karotenoid och baterioklorofyll i sina kromatoforpartiklar. Dessa molekyler hjälper till att absorbera ljus och omvandla det till energi och ger den också sin distinkta lilaröda färg under anaeroba förhållanden. R. rubrum är färglös under aeroba förhållanden.
Och även om fotosyntesen är aktiv under aeroba förhållanden, undertrycks den i allmänhet i närvaro av O2. Svavel är en viktig biprodukt av fotosyntesen, inte O2. R. rubrum kan växa heterotroft eller autotroft när den är fotosyntetisk. Till skillnad från många växter innehåller R. rubrum inget klorofyll a (absorptionsspektrum 430-662 nm). Den innehåller dock klorofyll b (absorptionsspektrum 660-680 nm) och bakterioklorofyller (800-925 nm). Detta gör att den kan utnyttja mer energi från de elektromagnetiska spektrumen. R. rubrum oxiderar också kolmonoxid (CO) med vätgas som slutprodukt på vägen och kan använda sulfid i låga koncentrationer som elektrondonator vid reduktion av koldioxid. Dessutom är den en kvävefixerande bakterie; den använder nitrogenas för att omvandla atmosfärisk kvävgas till ammoniak (Munk et al, 2011).
Det finns flera tillämpningar av R. rubrum inom bioteknikområdet. Den är ett modellsystem för omvandling av ljus till kemisk energi och för sina kvävefixeringsvägar. Den är också föremål för studier av strålningsresistens. Den kan också användas på flera sätt för konsumtion. Proteobakterierna kan vara en källa till djurfoder och jordbruksgödsel. En annan viktig roll inom forskningen omfattar produktion av vitaminer. Den undersöks också för sin produktion av biologisk plast från prekursorer av polyhydroxibutronsyra. R. rubrum kan också vara en bidragande orsak till biologiska vätgasbränslen, främst genom sin utveckling av enzymet nitrogenas.
Genomstruktur
Genomsekvenseringen av genomet är klar. Den består av en cirkulär kromosom med 4 352 825 baspar, varav 65 % är guanin-cystosinpar. Det finns också en plasmid med 53 732 baspar med 60 % guanin-cytosin. Totalt finns det 3 850 proteinkodande gener och 83 RNA-gener. Av dessa gener är 6,9 % transkriptionsrelaterade, 4,6 % translation, ribosomstruktur och biosyntes, 4,0 % replikation, rekombination och reparation, 7,9 % signaltransduktionsmekanismer, 5,9 % cellväggs- och membranbiogenes, 6,6 % energiproduktion och -omvandling, 5,0 % kolhydrattransport och -metabolism, 9.9 % transport och metabolism av aminosyror, 4,7 % transport och metabolism av koenzymer, 3,7 % transport och metabolism av lipider och 6,5 % transport och metabolism av oorganiska joner (Munk et al, 2011).
Ekologi
På grund av att Rhodospirillum rubrum kan växa både aerobt och anaerobt kan den leva under många olika förhållanden. R. rubrum finns i många naturliga vattenmiljöer såsom dammar, sjöar, vattendrag och stillastående vatten (Reslewic et. al, 2005). R. rubrum hittas också ofta i lera och avloppsvatten (Brock et al., 2000). Studier har visat att R. rubrum kan göra stora förändringar i sin kemiska sammansättning för att anpassa sig till olika miljöer (Cohen-Bazire och Kunisawa, 1963). R. rubrum föredrar att växa i livsmiljöer med ett pH på 6,8-7,2 (Bergey och Holt, 1994). Studier har också visat att R. rubrum har en optimal tillväxttemperatur på 22-35 grader Celcius (Weaver, 1971).
Cellstruktur och ämnesomsättning
R. rubrum är en mångsidig organism som kan få energi genom en mängd olika mekanismer. Respiration och fotosyntetiska mekanismer existerar tillsammans och deras aktivitet är beroende av närvaron av ljus och energi. R. rubrum kan växa i mörka kemotropiska miljöer med närvaro av syre eller kan växa i en fototropisk miljö utan syre.
Fotosyntesen hos R. rubrum börjar med donationen av ett väte från ett organiskt substrat till ett oxiderande substrat. Bakterioklorofyll och kartenoider, de fotoreaktiva pigmenten som finns i cellmembranet, är bundna till kromatophorer (Schachman, Pardee och Stanier, 1952). Chromatophorerna är tillplattade skivor som innehåller kolinfosfolipider, kardiolipin och galaktosyldiglycerider (Benson, Wintermans och Wiser, 1959; Benson 1961). Dessutom innehåller kromatofoner en komplett elektrontransportkedja som innehåller olika cytokromer, flavin och pyridinnukleotider (Newton och Newton, 1957; Hulcher och Conti, 1960; Kamen, 1961). Cellmembranet innehåller också maskiner för ATP-syntes, inklusive ett ATP-syntetas.
I närvaro av syre kan R. rubrum andas aerobt med hjälp av en traditionell elektrontransportkedja med NAD+/NADH som den primära elektronbäraren (Keister och Minton, 1969). Bland elektrontransportproteinerna finns cytokrom C428 (Chance och Smith, 1955). Aerob respiration hämmas dock av fotosyntesen (Oelze och Weaver, 1971).
R. rubrum kan också anaerobt andas. Dess icke-oxygena terminala elektronacceptorer kan inkludera dimetylsulfoxid och trimetylaminoxid. Närvaron av dessa elektronacceptorer gör det möjligt för substrat som succinat, malat och acetat att stödja tillväxten av R. rubrum. Dessa acceptorer är dock endast cirka 33-41 % lika effektiva som syre när det gäller energihushållning (Schultz och Weaver, 1982).
Dessutom har R. rubrum den unika förmågan att oxidera kolmonoxid med hjälp av kolmonoxiddehydrogenas. Denna oxidationsväg slutar med reduktion och väte och produktion av vätgas.
Och utan en terminal elektronacceptor fullbordar R. rubrum blandsyrafermentering. De viktigaste produkterna från pyruvatjäsningen är acetat, format, koldioxid och väte. I närvaro av bikarbonatjonen kan fruktos fermenteras. Slutprodukterna för denna väg är desamma som för pyruvat men med tillägg av succinat och propionat (Schultz och Weaver 1982).
Bortsett från förmågan att fixera koldioxid (Schon och Biedermann, 1972) är en identifierande anabolisk egenskap hos R. rubrum dess förmåga att fixera kväve. Under mörka förhållanden med närvaro av fruktos använder R. rubrum nitrogenas för att fixera kvävgas till ammonium. Den innehåller både Fe-Mo och Fe-kvävgas.
Patologi
R. rubrum har inte visat sig infektera människor eller djur.
Aktuell forskning
Strålningsresistens
Rhodospirillum rubrum har nyligen gett insikt om strålningsresistens. Felice Mastroleo et al har i Insight into the radiotolerance of the life support bacterium Rhodospirillum rubrum S1H by means of phenotypic and transcriptomic methods (2010) karakteriserat R. rubrums strålningstolerans.
Genom att utsätta bakterien för subdödliga doser kobolt-60 gammastrålar, en typ av joniserande strålning, karakteriserades R. rubrums markörgener som är specifika för joniserande strålning i organismen. Dessa gener ökade i uttryck i takt med att ”återhämtningstiden” förflöt efter en given gammastrålning. Analysen av genuttryck hos R. rubrum efter gammastrålning skapar potential för ytterligare mikrobiologisk forskning som kan leda till framsteg inom tillämpad strålningsresistens i framtiden.
Biologiskt nedbrytbara plaster
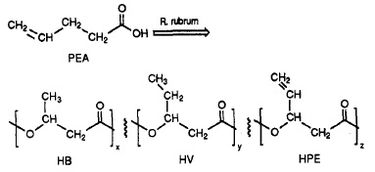
R. rubrum har visat sig producera en klass av biologiskt nedbrytbara plaster, polybetahydroxyalkanoater (PHA). Tillämpningarna av denna forskning kan lösa många problem som orsakas av syntetiska, icke nedbrytbara plaster som har påverkat miljön negativt. Ett forskningsexperiment som utfördes av Herbert. W. Ulmer et al. gjordes en krönika om produktion av PHA:er som innehåller 3HV-enheter av Rhodospriillum rubrum (1993). Ulmer använde 4-pentensyra (PEA) som tillväxtmedium för R. rubrum. När bakterien placerades under ”PHA-producerande förhållanden” syntetiserades PHA:er som innehöll tre upprepade underenheter (not: R = substituentgrupp): HB (R = metyl), HV (R = etyl), HPE (R = vinyl) (se figur 1).
Syntesen av funktionella, biologiskt nedbrytbara plaster med hjälp av bakterier har potential att minska den globala föroreningen och bränsleförbrukningen från plastfabriker. R. rubrum kan användas som ett mikrobiologiskt verktyg för att producera miljövänliga biopolymerer.
Renoverbar energi
Den ökande oron för miljöföroreningar från förbränning av fossila bränslen har lett till att forskningen om Rhodospirillum rubrum har ökat. Genom att använda R. rubrum som en källa för vätgas som ska användas i förnybara vätgasbränslen åtgärdas flera problem som skapats av förbrukningen av fossila bränslen. Vätgas oxideras till vatten som en förbränningsprodukt, till skillnad från fossila bränslen som skapar föroreningar som CO, CO2, SOx och NOx. R. rubrum kan producera vätgas genom att utnyttja följande ”vatten-gasförskjutningsreaktion” (Najafpour et al., 2004):
CO + H2O → H2 + CO2
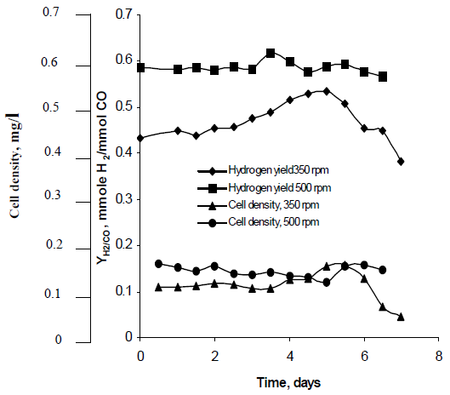
Najafpour observerade hur omrörning, som skapades av turbinhjul som ställdes in med hastigheterna 350 varv per minut samt 500 varv per minut, påverkade vätgasproduktionshastigheterna hos R. rubrum. Hans forskning visade att vätgasbränsle har potential att bromsa den globala uppvärmningen och erbjuda en miljövänlig teknisk lösning för bränsleproduktion och bränsleförbrukning. Hans studie visade att ”högre omrörningshastighet gav högre vätgas och högre celltäthet” hos R. rubrum (se figur 2).
I en annan mikrobiologisk studie, utförd av Matthew Ross Melnicki, fann man att R. rubrum ger potential för förnybar energi, såsom vätgasbränsle, via vätgassyntes. Dess biologiska vägar kan producera vätgasnivåer med hög hastighet när den samodlas med Chlamydomonas reinhardtii, en algart som R. rubrum kan växa harmoniskt med. Melnicki föreslog ett okonventionellt tillvägagångssätt för samodling för att syntetisera vätgas som gav ökade nivåer av vätgassyntes som kanske inte kan uppnås om samodlingen är frikopplad. C. reinhardtii gynnades vid högre ljusintensitet, medan R. ruburm var mer produktiv vid lägre ljusintensitet, vilket belyser fördelarna med deras samodling.
Med hjälp av en fed-batch-teknik för odling av R. rubrum fann man att inokulerade kulturer producerade vätgas under dess exponentiella tillväxtfas, och vätgassyntesen fortsatte att blomstra kontinuerligt i 70 timmar. När tillväxten upphörde tillsatte Melnicki succinat, som fungerar som en elektrondonator i den metaboliska vägen, och vätgasutvecklingen återupptogs även om bakterierna nu inte längre växte. Det är dock viktigt att notera att vid kontinuerlig tillsats av succinat fann man minskad utvecklingshastighet av väte. När ytterligare tillväxtmedium tillsattes ökade vätgasproduktionshastigheten igen.
Melnicki konstaterade också att ”även om tillväxt inte krävs för vätgasproduktion, fastställer detta arbete behovet av celltillväxt för att upprätthålla maximala hastigheter, vilket tyder på industriell lämplighet för en halvkontinuerlig odlingsstrategi” (Melnicki, 2009).
Cool Factor
Genomsekvensen av Rhodospirillum rubrum är bara den andra av Rhodospirillum-släktet som har publicerats, efter R. centenum (Munk et al, 2011).

En intressant aspekt av Rhodospirillum rubrum är dess förmåga att ändra färg under olika förhållanden. Den är en anoxygen fototrof, men producerar inte extracellulärt syre. Istället producerar den elementärt svavel under ljusskörden. Detta beror på att R. rubrum saknar light harvesting complex 2 (Munk et al, 2011). Bakterien använder bakterioklorofyll, vilket gör att den kan absorbera våglängder av ljus som växter inte utnyttjar. Bakterioklorofyll absorberar våglängder på 800-925 nm.
Under aeroba förhållanden är den fotosyntetiska aktiviteten hos R. rubrum genetiskt undertryckt och bakterien blir färglös. Under anaeroba förhållanden aktiveras dock fotosyntesen och bakterien får en tydlig lilaröd färg (se bild 5). Detta beror på aktivitet i kromatoforerna, som finns i membranet (Munk et al, 2011). Kromatoforerna innehåller bakteriens bakterieklorofyllpigment och genomgår absorptionsförändringar vid aktivering av fotosyntesen.
Benson, A. ”Chloroplast lipid metabolism”. I Symposium nr. VI. 5th international Congress on Biochemistry. 1961. Oxford: Pergamon Press. (I tryck.)
Benson, A., Wintermans, J. och Wiser, R. ”Chloroplast lipids as carbohydrate reservoirs”. Plant Physiology. 1959. Volume 34. p. 815.
Bergey, D. and Holt, J. Bergey’s manual of determinative bacteriology. Nionde upplagan. Baltimore. Lippincott Williams and Wilkins. 1994. s. 359.
Brock, T.D., M.T. Madigan, J.M. Martinko och J Parker. Biology of Microorganisms. 9th Edition. Upper Saddle River: Prentice Hall, 2000. s. 459.
Chance, B. och Smith, L. ”Andningspigment hos Rhodospirillum rubrum.” Natur. Volym 174. s. 803-806.
Cohen-Bazire, G. och Kunisawa, R. ”The Fine Structure of Rhodospirillum Rubrum.” Journal of Cell Biology. 1963. Volume 16. p. 401-419.
Hulcher, M. and Conti, S. ”Cytochronies in chlorophyll-containing particles of Chromatium and Chlorobium thiosuphatophilum”. Biochemistry and Biophysics Research Communications. 1960. Volume 3. s. 497.
Kamen, M. ”The Haem Proteins of Photosynthetic Tissues”. In Symposium no. VI. 5th international Congress on Biokemi. 1961. Oxford: Pergamon Press. (I tryck.)
Keister, D. and Minton, N. ”Energy-linked reactions in photosynthetic bacteria. 3. Further studies on energy-linked nicotinamide-adenine dinucleotide reduction by Rhodospirillum rubrum chromatophores”. Biochemistry. 1969. Volume 8. p. 167-173.
Mastroleo, F., Monsieurs, P., and Leys, N. ”Insight into the radiotolerance of the life support bacterium Rhodospirillum rubrum S1H by means of phenotypic and transcriptomic methods”. COSPAR:s vetenskapliga församling. 2010. s. 4.
Melnicki, M. R. ”Modes of hydrogen production in the photosynthetic bacterium, Rhodosphirillum rubrum”. University of California, Berkeley. 2009.
Munk, C., Copeland, A., Lucas, C., Lapidus, A., et al. ”Complete genome sequence of Rhodospirillum rubrum type strain (S1T)”. Standards in Genomic Sciences. 2011. Volume 4. pp. 293-302.
Najafpour, G., Ismail, K., Younesi, H., Mohamed, A. R., and Kamaruddin, A. H. ”Hydrogen as clean fuel via continuous fermentation by anaerobic photosynthetic bacteria, Rhodospirillum rubrum”. African Journal of Biotechnology. 2004. Volume 3. pp. 503-507.
Newton, J. and Newton, G. ”Composition of the photoactive subcellular particles from Chromatium”. Archives of Biochemistry and Biophysics. 1957. Volume 71. p. 250.
Oelze, J. and Weaver, P. ”The adjustment of photosynthetically grown cells of Rhodospirillum rubrum to aerobic light conditions”. The Archives of Microbiology. 1982. Volume 79. p. 108-121.
Reslewic, S., Shigou, Z., Place, M., Zhang, Y., Briska A., Goldstein, S., Churas, C., Runnheim, R., Forrest, D., Lim, A., Lapidus, A., Han, C., Roberts, G., and Schwartz, D. ”Whole-Genome Shotgun Optical Mapping of Rhodospirillum rubrum”. Tillämpad och miljömässig mikrobiologi. 2005. Volym 71. s. 5511-5522.
Schachman, H., Pardee, A. och Stanier, R. ”Studies on the macromolecular organization of microbial cells”. Archives of Biochemistry and Biophysics. 1952. Volume 38. p. 245-260.
Schön, G. and Biedermann, M. ”Synthesis of volatile acids by fermentation of pyruvate and fructose in anaerobic dark cultures of Rhodospirillum rubrum”. Archiv fur Mikrobiologie.1972. Volume 85. p. 77-90.
Schultz, J. and Weaver, P. ”Fermentation and anaerobic respiration by Rhodospirillum rubrum and Rhodopseudomonas capsulate”. The Journal of Bacteriology. 1982. Volume 149. p. 181-190.
Ulmer, H., Gross, R., Posada, M., Weisbach, P., Fuller, R. C., and Lenz, R. ”Bacterial production of poly(.beta.-hydroxyalkanoates) containing unsaturated repeating units by Rhodospirillum rubrum”. Makromolekyler. 1994. Volume 27. p. 1675-1679.
Weaver, P. ”Temperature-Sensitive Mutations of the Photosynthetic Apparatus of Rhodospirillum rubrum”. Proceedings of the National Academy of Sciences of the United States of America. 1971. Volume 68. p. 136-138.
Redigerat av studenter till Dr. Iris Keren: Allison Almeida, Matthew Cozzolino, Trung Nguyen och Jessica Wong.
NEUF2011