Kvävebaser, viktiga komponenter i nukleotider, är organiska molekyler och kallas så eftersom de innehåller kol och kväve. De är baser eftersom de innehåller en aminogrupp som har potential att binda ett extra väte och därmed minskar vätejonkoncentrationen i sin omgivning, vilket gör den mer basisk. Varje nukleotid i DNA innehåller en av fyra möjliga kvävebaser: adenin (A), guanin (G), cytosin (C) och tymin (T). RNA-nukleotider innehåller också en av fyra möjliga baser: adenin, guanin, cytosin och uracil (U) i stället för tymin.
Adenin och guanin klassificeras som puriner. Den primära strukturen hos ett purin är två kol-stickväte ringar. Cytosin, tymin och uracil klassificeras som pyrimidiner som har en enda kolväte-kväve-ring som primärstruktur (figur 1). Var och en av dessa grundläggande kol-kväve-ringar har olika funktionella grupper knutna till sig. I molekylärbiologisk förkortning kallas kvävebaserna helt enkelt för symbolerna A, T, G, C och U. DNA innehåller A, T, G och C medan RNA innehåller A, U, G och C.
Pentosockret i DNA är deoxyribose och i RNA är sockret ribose (figur 1). Skillnaden mellan sockerarterna är att det finns en hydroxylgrupp på det andra kolet i ribose och väte på det andra kolet i deoxyribose. Kolatomerna i sockermolekylen är numrerade som 1′, 2′, 3′, 4′ och 5′ (1′ betyder ”ett primtal”). Fosfatresterna är bundna till hydroxylgruppen på 5′-kolvret i ett socker och hydroxylgruppen på 3′-kolvret i sockret i nästa nukleotid, vilket bildar en 5′-3′-fosfodiesterbindning. Fosfodiesterlänken bildas inte genom en enkel dehydreringsreaktion som andra länkar som förbinder monomerer i makromolekyler: för att den ska bildas måste två fosfatgrupper avlägsnas. En polynukleotid kan ha tusentals sådana fosfodiesterlänkar.
DNA Double-Helix Structure
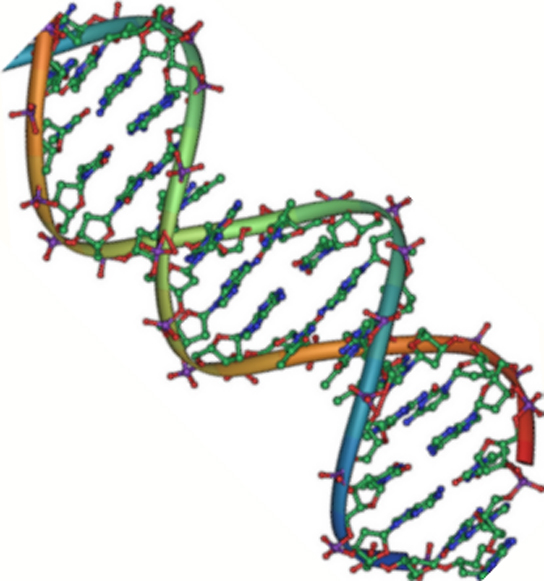
Figur 2. DNA är en antiparallell dubbelhelix. Fosfatryggen (de krökta linjerna) är på utsidan och baserna är på insidan. Varje bas interagerar med en bas från den motsatta strängen. (kredit: Jerome Walker/Dennis Myts)
DNA har en dubbelhelixstruktur (figur 2). Sockret och fosfatet ligger på utsidan av spiralen och bildar DNA:s ryggrad. Kvävebaserna är staplade på insidan, likt trappstegen i en trappa, i par; paren är bundna till varandra genom vätebindningar. Varje baspar i dubbelhelixen är separerat från nästa baspar med 0,34 nm.
De två strängarna i helixen löper i motsatt riktning, vilket innebär att den ena strängens 5′-karbonände är vänd mot den motsvarande strängens 3′-karbonände. (Detta kallas antiparallell orientering och är viktigt för DNA-replikation och i många nukleinsyrainteraktioner.)
Endast vissa typer av basparning är tillåtna. Till exempel kan ett visst purin bara para sig med ett visst pyrimidin. Detta innebär att A kan para sig med T och G kan para sig med C, vilket visas i figur 3. Detta är känt som baskomplementärregeln. Med andra ord är DNA-strängarna komplementära till varandra. Om sekvensen i den ena strängen är AATTGGCC skulle den komplementära strängen ha sekvensen TTAACCGG. Under DNA-replikationen kopieras varje sträng, vilket resulterar i en dotter-DNA-dubbelhelix som innehåller en föräldra-DNA-sträng och en nysyntetiserad sträng.
Praktikfråga
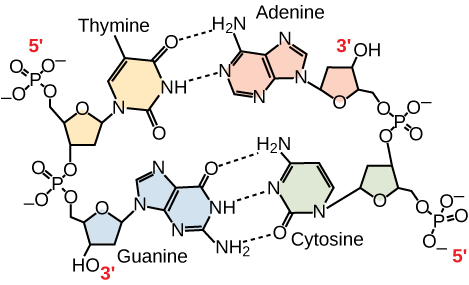
Figur 3. I en dubbelsträngad DNA-molekyl löper de två strängarna antiparallel till varandra så att den ena strängen löper 5′ till 3′ och den andra 3′ till 5′. Fosfatryggen ligger på utsidan och baserna ligger i mitten. Adenin bildar vätebindningar (eller baspar) med tymin och guanin baspar med cytosin.
En mutation inträffar och cytosin ersätts med adenin. Vilken inverkan tror du att detta kommer att ha på DNA-strukturen?
RNA
Ribonukleinsyra, eller RNA, är huvudsakligen involverad i proteinsyntesen under ledning av DNA. RNA är vanligtvis enkelsträngat och består av ribonukleotider som är sammankopplade med fosfodiesterbindningar. En ribonukleotid i RNA-kedjan innehåller ribose (pentosocker), en av de fyra kvävebaserna (A, U, G och C) och fosfatgruppen.
Det finns fyra huvudtyper av RNA: budbärar-RNA (mRNA), ribosomalt RNA (rRNA), överförings-RNA (tRNA) och mikroRNA (miRNA). Det första, mRNA, bär budskapet från DNA, som styr alla cellaktiviteter i en cell. Om en cell kräver att ett visst protein syntetiseras, aktiveras genen för denna produkt och budbärar-RNA syntetiseras i cellkärnan. RNA-bassekvensen är komplementär till kodningssekvensen i det DNA från vilket den har kopierats. I RNA saknas dock basen T och i stället finns U. Om DNA-strängen har sekvensen AATTGCGC är sekvensen för det komplementära RNA:t UUAACGCG. I cytoplasman interagerar mRNA med ribosomer och andra cellulära maskiner (figur 4).
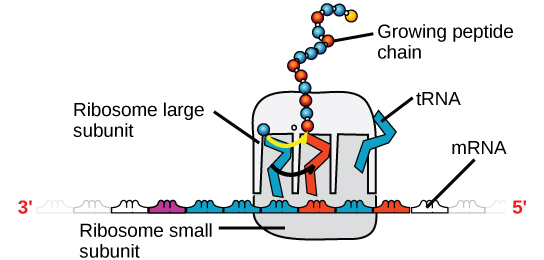
Figur 4. En ribosom har två delar: en stor underenhet och en liten underenhet. Det mRNA sitter mellan de två subenheterna. En tRNA-molekyl känner igen en kodon på mRNA, binder till den genom komplementär basparning och lägger till rätt aminosyra till den växande peptidkedjan.
Det mRNA avläses i uppsättningar av tre baser som kallas kodoner. Varje kodon kodar för en enda aminosyra. På detta sätt avläses mRNA och proteinprodukten tillverkas. Ribosomalt RNA (rRNA) är en viktig beståndsdel i de ribosomer på vilka mRNA binds. RRNA ser till att mRNA och ribosomerna är rätt inriktade; ribosomens rRNA har också en enzymatisk aktivitet (peptidyltransferas) och katalyserar bildandet av peptidbindningar mellan två inriktade aminosyror. Transfer-RNA (tRNA) är en av de minsta av de fyra typerna av RNA, vanligtvis 70-90 nukleotider lång. Det transporterar rätt aminosyra till platsen för proteinsyntesen. Det är basparningen mellan tRNA och mRNA som gör att rätt aminosyra kan infogas i polypeptidkedjan. mikroRNA är de minsta RNA-molekylerna och deras roll innebär reglering av genuttrycket genom att störa uttrycket av vissa mRNA-meddelanden.