Le basi azotate, importanti componenti dei nucleotidi, sono molecole organiche e sono così chiamate perché contengono carbonio e azoto. Sono basi perché contengono un gruppo amminico che ha il potenziale di legare un idrogeno extra, e quindi, diminuisce la concentrazione di ioni idrogeno nel suo ambiente, rendendolo più basico. Ogni nucleotide nel DNA contiene una delle quattro possibili basi azotate: adenina (A), guanina (G) citosina (C) e timina (T). Anche i nucleotidi dell’RNA contengono una delle quattro possibili basi: adenina, guanina, citosina e uracile (U) piuttosto che timina.
Adenina e guanina sono classificate come purine. La struttura primaria di una purina è costituita da due anelli di carbonio-azoto. La citosina, la timina e l’uracile sono classificati come pirimidine che hanno un singolo anello di carbonio-azoto come struttura primaria (Figura 1). Ognuno di questi anelli di carbonio-azoto di base ha diversi gruppi funzionali attaccati ad esso. In biologia molecolare, le basi azotate sono semplicemente conosciute con i loro simboli A, T, G, C e U. Il DNA contiene A, T, G e C, mentre l’RNA contiene A, U, G e C.
Lo zucchero pentoso nel DNA è il deossiribosio, e nell’RNA, lo zucchero è il ribosio (Figura 1). La differenza tra gli zuccheri è la presenza del gruppo idrossile sul secondo carbonio del ribosio e dell’idrogeno sul secondo carbonio del desossiribosio. Gli atomi di carbonio della molecola di zucchero sono numerati come 1′, 2′, 3′, 4′, e 5′ (1′ si legge come “un primo”). Il residuo di fosfato è attaccato al gruppo idrossile del carbonio 5′ di uno zucchero e al gruppo idrossile del carbonio 3′ dello zucchero del nucleotide successivo, che forma un legame fosfodiestere 5′-3′. Il legame fosfodiestere non si forma per semplice reazione di disidratazione come gli altri legami che collegano i monomeri nelle macromolecole: la sua formazione comporta la rimozione di due gruppi fosfato. Un polinucleotide può avere migliaia di tali legami fosfodiesteri.
Struttura del doppio elice del DNA
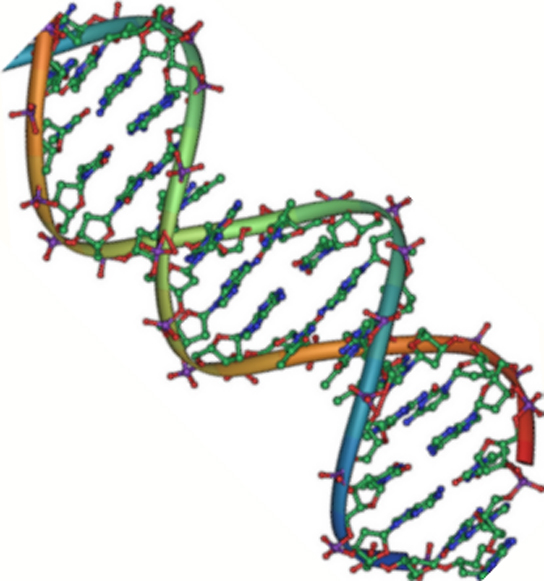
Figura 2. Il DNA è una doppia elica antiparallela. La spina dorsale di fosfato (le linee curve) è all’esterno, e le basi sono all’interno. Ogni base interagisce con una base del filamento opposto. (credit: Jerome Walker/Dennis Myts)
Il DNA ha una struttura a doppia elica (Figura 2). Lo zucchero e il fosfato si trovano all’esterno dell’elica, formando la spina dorsale del DNA. Le basi azotate sono impilate all’interno, come i gradini di una scala, a coppie; le coppie sono legate tra loro da legami idrogeno. Ogni coppia di basi nella doppia elica è separata dalla coppia di basi successiva da 0,34 nm.
I due filamenti dell’elica corrono in direzioni opposte, il che significa che l’estremità di carbonio 5′ di un filamento si trova di fronte all’estremità di carbonio 3′ del filamento corrispondente. (Questo è indicato come orientamento antiparallelo ed è importante per la replicazione del DNA e in molte interazioni dell’acido nucleico.)
Solo alcuni tipi di accoppiamento di basi sono permessi. Per esempio, una certa purina può accoppiarsi solo con una certa pirimidina. Questo significa che A può accoppiarsi con T, e G può accoppiarsi con C, come mostrato nella Figura 3. Questa è conosciuta come la regola della complementarietà delle basi. In altre parole, i filamenti di DNA sono complementari tra loro. Se la sequenza di un filamento è AATTGGCC, il filamento complementare avrebbe la sequenza TTAACCGG. Durante la replicazione del DNA, ogni filamento viene copiato, ottenendo una doppia elica di DNA figlia contenente un filamento di DNA parentale e un filamento appena sintetizzato.
Domanda pratica
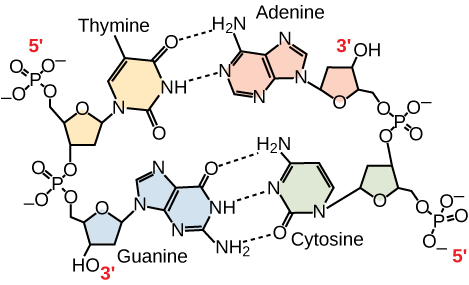
Figura 3. In una molecola di DNA a doppio filamento, i due filamenti corrono antiparalleli l’uno all’altro in modo che un filamento corra da 5′ a 3′ e l’altro da 3′ a 5′. La spina dorsale del fosfato si trova all’esterno e le basi sono al centro. L’adenina forma legami idrogeno (o coppie di basi) con la timina, e la guanina forma coppie di basi con la citosina.
Si verifica una mutazione, e la citosina viene sostituita dall’adenina. Che impatto pensi che questo avrà sulla struttura del DNA?
RNA
L’acido ramonucleico, o RNA, è principalmente coinvolto nel processo di sintesi delle proteine sotto la direzione del DNA. L’RNA è di solito a filamento singolo ed è composto da ribonucleotidi legati da legami fosfodiesteri. Un ribonucleotide nella catena dell’RNA contiene ribosio (lo zucchero pentoso), una delle quattro basi azotate (A, U, G e C) e il gruppo fosfato.
Ci sono quattro tipi principali di RNA: RNA messaggero (mRNA), RNA ribosomiale (rRNA), RNA di trasferimento (tRNA) e microRNA (miRNA). Il primo, l’mRNA, porta il messaggio del DNA, che controlla tutte le attività cellulari di una cellula. Se una cellula richiede la sintesi di una certa proteina, il gene per questo prodotto viene acceso e l’RNA messaggero viene sintetizzato nel nucleo. La sequenza di base dell’RNA è complementare alla sequenza codificante del DNA da cui è stata copiata. Tuttavia, nell’RNA, la base T è assente e al suo posto è presente la U. Se il filamento di DNA ha una sequenza AATTGCGC, la sequenza dell’RNA complementare è UUAACGCG. Nel citoplasma, l’mRNA interagisce con i ribosomi e altri macchinari cellulari (Figura 4).
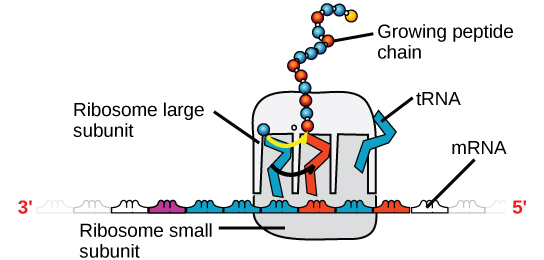
Figura 4. Un ribosoma ha due parti: una subunità grande e una subunità piccola. L’mRNA si trova tra le due subunità. Una molecola di tRNA riconosce un codone sull’mRNA, si lega ad esso tramite un accoppiamento di basi complementari e aggiunge l’amminoacido corretto alla catena peptidica in crescita.
L’mRNA viene letto in serie di tre basi note come codoni. Ogni codone codifica per un singolo amminoacido. In questo modo, l’mRNA viene letto e viene realizzato il prodotto proteico. L’RNA ribosomiale (rRNA) è uno dei principali costituenti dei ribosomi su cui si lega l’mRNA. L’rRNA assicura il corretto allineamento dell’mRNA e dei ribosomi; l’rRNA del ribosoma ha anche un’attività enzimatica (peptidil transferasi) e catalizza la formazione dei legami peptidici tra due amminoacidi allineati. L’RNA di trasferimento (tRNA) è uno dei più piccoli dei quattro tipi di RNA, solitamente lungo 70-90 nucleotidi. Trasporta l’aminoacido corretto al sito di sintesi proteica. È l’accoppiamento di base tra il tRNA e l’mRNA che permette all’amminoacido corretto di essere inserito nella catena polipeptidica. I microRNA sono le molecole di RNA più piccole e il loro ruolo riguarda la regolazione dell’espressione genica interferendo con l’espressione di certi messaggi di mRNA.