A mikrobiális biorégió oldala a Rhodospirillum rubrum nemzetségről
Klasszifikáció
Felsőbb rendű taxonok
Királyság: Baktériumok
Törzs: Proteobaktériumok
Osztály: Osztály: Alphaproteobacteria
Rend: Rhodospirillales
Family: Rhodospirillaceae
Genus: Rhodospirillum
Fajok
Rhodospirillum rubrum
Megnevezés és jelentőség
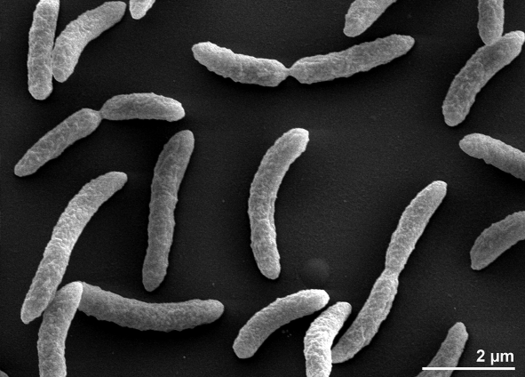
A Rhodospirillum rubrum Gram-negatív, mezofil proteobaktérium. Optimális növekedési hőmérséklete 25-30 Celsius-fok. Többrétegű külső burokkal rendelkezik, amely sejtfalában többnyire telítetlen, de néhány telített zsírt tartalmaz. A R. rubrum spirilla, azaz spirál alakú. Polárisan flagellált, ezért mozgékony. Hossza 3-10 um, szélessége 0,8-1,0 um.
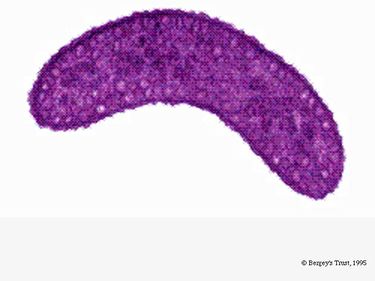
A R. rubrum fakultatív anaerob. Oxigén jelenlététől függően alkoholos erjedésen vagy aerob légzésen mehet keresztül. Fotoszintézisre is képes, kromatofór részecskéiben karotinoidot és baterioklorofillt tartalmaz. Ezek a molekulák segítenek a fény elnyelésében és energiává alakításában, valamint anaerob körülmények között jellegzetes lilásvörös színt kölcsönöznek neki. A R. rubrum aerob körülmények között színtelen.
A fotoszintézis aerob körülmények között ugyan aktív, de O2 jelenlétében általában elnyomódik. A fotoszintézis fő mellékterméke a kén, nem az O2. A R. rubrum fotoszintézis esetén heterotróf vagy autotróf módon növekedhet. Sok növénytől eltérően az R. rubrum nem tartalmaz klorofill a-t (abszorpciós spektrum 430-662 nm). Tartalmaz azonban b-klorofillt (abszorpciós spektrum 660-680 nm) és baktérium-klorofillt (800-925 nm). Ez lehetővé teszi, hogy több energiát hasznosíthasson az elektromágneses spektrumból. A R. rubrum oxidálja a szén-monoxidot (CO) is, amelynek végterméke hidrogéngáz, és alacsony koncentrációban szulfidot is képes elektrondonorként használni a szén-dioxid redukciójában. Emellett nitrogénkötő baktérium; nitrogenáz segítségével a légköri nitrogéngázt ammóniává alakítja (Munk et al, 2011).
A R. rubrumnak számos alkalmazása van a biotechnológia területén. A fény kémiai energiává történő átalakításának és nitrogénkötő útvonalainak modellrendszere. Sugárrezisztencia-vizsgálatok tárgyát is képezi. Többféleképpen felhasználható fogyasztásra is. A proteobaktérium állati takarmány és mezőgazdasági műtrágya forrása lehet. A kutatásban fontos szerepet játszik még a vitaminok előállítása. A poli-hidroxi-vajsav prekurzoraiból biológiai műanyag előállítását is kutatják. A R. rubrum a biológiai hidrogén-üzemanyagokban is közreműködhet, főként a nitrogenáz enzim fejlesztése révén.
Genomszerkezet
A genom szekvenálása befejeződött. Egy 4 352 825 bázispárból álló körkörös kromoszómából áll, amelynek 65%-a guanin-cisztozin-pár. Van egy plazmid is 53 732 bázispárral, amelynek 60%-a guanin-citozin. Összesen 3850 fehérjét kódoló gén és 83 RNS-gén van. Ezen gének 6,9%-a transzkripcióval kapcsolatos; 4,6%-a transzláció, riboszóma szerkezet és bioszintézis; 4,0%-a replikáció, rekombináció és javítás; 7,9%-a jelátviteli mechanizmusok; 5,9%-a sejtfal és membrán biogenezis; 6,6%-a energiatermelés és -átalakítás; 5,0%-a szénhidrát-transzport és -anyagcsere; 9. A gének közül 6,9%-nak a transzkripcióhoz van köze.9% aminosav transzport és anyagcsere; 4,7% koenzim transzport és anyagcsere; 3,7% lipid transzport és anyagcsere; és 6,5% szervetlen ion transzport és anyagcsere(Munk et al, 2011).
Ökológia
A Rhodospirillum rubrum aerob és anaerob módon egyaránt képes növekedni, így a legkülönbözőbb körülmények között képes megélni. Az R. rubrum számos természetes vízi környezetben, például tavakban, tavakban, patakokban és állóvizekben megtalálható (Reslewic et. al, 2005). A R. rubrum gyakran megtalálható iszapban és szennyvízben is (Brock et al, 2000). Vizsgálatok kimutatták, hogy a R. rubrum képes nagymértékben megváltoztatni kémiai összetételét, hogy alkalmazkodjon a különböző környezetekhez (Cohen-Bazire és Kunisawa, 1963). A R. rubrum a 6,8-7,2 pH-értékű élőhelyeket kedveli (Bergey és Holt, 1994). A vizsgálatok azt is kimutatták, hogy a R. rubrum optimális növekedési hőmérséklete 22-35 Celsius-fok (Weaver, 1971).
Cellaszerkezet és anyagcsere
A R. rubrum sokoldalú szervezet, amely többféle mechanizmuson keresztül képes energiát nyerni. A légzési és fotoszintetikus mechanizmusok együtt léteznek, és aktivitásuk a fény és az energia jelenlététől függ. A R. rubrum képes sötét kemotropikus környezetben oxigén jelenlétében, vagy oxigén nélküli fototropikus környezetben növekedni.
A fotoszintézis a R. rubrumban azzal kezdődik, hogy egy szerves szubsztrátból hidrogént adományoz egy oxidáló szubsztrátnak. A baktérium-klorofill és a kartenoidok, a sejtmembránban található fotoreaktív pigmentek kromatofórokhoz kötődnek (Schachman, Pardee és Stanier, 1952). A kromatofórák lapított korongok, amelyek kolin-foszfolipideket, kardiolipint és galaktozil-diglicerideket tartalmaznak (Benson, Wintermans és Wiser, 1959; Benson 1961). Ezenkívül a kromatofórák teljes elektrontranszportláncot tartalmaznak, amely különböző citokrómokat, flavint és piridin-nukleotidokat tartalmaz (Newton és Newton, 1957; Hulcher és Conti, 1960; Kamen, 1961). A sejtmembrán az ATP-szintézis gépezetét is tartalmazza, beleértve egy ATP-szintetázt.
A R. rubrum oxigén jelenlétében aerob módon képes lélegezni egy hagyományos elektrontranszportlánc segítségével, amelyben a NAD+/NADH az elsődleges elektronhordozó (Keister és Minton, 1969). Az elektrontranszportfehérjék között van a citokróm C428 (Chance és Smith, 1955). Az aerob légzést azonban gátolja a fotoszintézis (Oelze és Weaver, 1971).
A R. rubrum anaerob légzésre is képes. Nem oxigénterminális elektronakceptorai közé tartozhat a dimetil-szulfoxid és a trimetilamin-oxid. Ezen elektronakceptorok jelenléte lehetővé teszi, hogy az olyan szubsztrátok, mint a szukcinát, a malát és az acetát támogassák a R. rubrum növekedését. Ezek az akceptorok azonban az energiamegtartás szempontjából csak körülbelül 33-41%-ban olyan hatékonyak, mint az oxigén (Schultz és Weaver, 1982).
A R. rubrum emellett egyedülálló módon képes a szénmonoxid oxidálására a szénmonoxid-dehidrogenáz segítségével. Ez az oxidációs út redukcióval és hidrogénnel, valamint hidrogéngáz termelésével végződik.
Terminális elektronakceptor nélkül az R. rubrum befejezi a vegyes savas fermentációt. A piruvátfermentáció fő termékei az acetát, a formiát, a szén-dioxid és a hidrogén. Bikarbonátion jelenlétében a fruktóz is képes fermentálni. Ennek az útvonalnak a végtermékei megegyeznek a piruvátéval, de szukcinát és propionát hozzáadásával (Schultz és Weaver 1982).
A szén-dioxid megkötésének képessége mellett (Schon és Biedermann, 1972) a R. rubrum egyik azonosító anabolikus tulajdonsága a nitrogén megkötésének képessége. Sötét körülmények között, fruktóz jelenlétében az R. rubrum nitrogenáz segítségével a nitrogéngázt ammóniává fixálja. Fe-Mo és csak Fe nitrogenázt is tartalmaz.
Patológia
R. rubrum nem fertőzött embert vagy állatot.
A jelenlegi kutatások
Sugárzásállóság
A Rhodospirillum rubrum a közelmúltban betekintést nyújtott a sugárzásállóságba. Felice Mastroleo és munkatársai az Insight into the radiotolerance of the life support bacterium Rhodospirillum rubrum S1H by means of phenotypic and transcriptomic methods (2010) című munkájukban jellemezték a R. rubrum sugárzástűrését.
A baktériumot szubletális dózisú Cobalt-60 gamma-sugárzásnak, egyfajta ionizáló sugárzásnak kitéve, jellemezték a szervezetben az R. rubrum ionizáló sugárzásra specifikus markergénjeit. Ezeknek a géneknek az expressziója az adott gammasugárzást követő “regenerációs idő” elteltével nőtt. A R. rubrum gamma-sugárzás utáni génexpressziójának elemzése lehetőséget teremt további mikrobiológiai kutatásokra, amelyek a jövőben az alkalmazott sugárrezisztencia terén előrelépésekhez vezethetnek.
Biodegradálható műanyagok
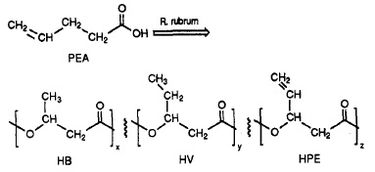
A R. rubrum a biológiailag lebomló műanyagok egy osztályát, a poli-béta-hidroxialkanoátokat (PHA-k) állítja elő. E kutatás alkalmazásai számos, a környezetet károsan terhelő szintetikus, nem lebomló műanyagok által okozott problémát oldhatnak meg. A Herbert által végzett kutatási kísérlet. W. Ulmer és munkatársai a Rhodospriillum rubrum (1993) által 3HV egységeket tartalmazó PHA-k előállítását dokumentálták. Ulmer 4-penténsavat (PEA) használt a R. rubrum növekedési közegeként. Amikor a baktériumot “PHA-termelő körülmények” közé helyezték, 3 ismétlődő alegységet tartalmazó PHA-kat szintetizáltak (megjegyzés: R = szubsztituenscsoport): HB (R = metil), HV (R = etil), HPE (R = vinil) (lásd az 1. ábrát).
A funkcionális, biológiailag lebomló műanyagok baktériumokkal történő szintézisével csökkenthető a globális szennyezés és a műanyaggyárak üzemanyag-ráfordítása. Az R. rubrum mikrobiológiai eszközként használható környezetbarát biopolimerek előállítására.
Újuló energia
A fosszilis tüzelőanyagok elégetéséből származó környezetszennyezéssel kapcsolatos növekvő aggodalom a Rhodospirillum rubrum kutatásának növekedéséhez vezetett. A R. rubrum megújuló hidrogénüzemanyagokban felhasználható hidrogénforrásként való felhasználása számos, a fosszilis tüzelőanyagok fogyasztása által okozott problémával foglalkozik. A hidrogén égéstermékként vízzé oxidálódik, ellentétben a fosszilis tüzelőanyagokkal, amelyek olyan szennyező anyagokat termelnek, mint a CO, CO2, SOx és NOx. Az R. rubrum a következő “víz-gáz eltolódási reakció” (Najafpour et al, 2004) segítségével képes hidrogént előállítani:
CO + H2O → H2 + CO2
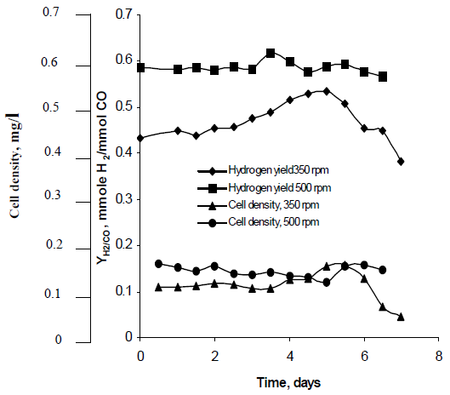
Najafpour kutatási módszerében megfigyelte, hogy a 350 rpm, valamint az 500 rpm sebességgel beállított turbinalapát-készletek által létrehozott keverés hogyan befolyásolja a R. rubrum hidrogéntermelésének sebességét. Kutatása megállapította, hogy a hidrogén üzemanyag képes megfékezni a globális felmelegedést, és környezetbarát technológiai megoldást kínál az üzemanyag-előállításra és -fogyasztásra. Vizsgálata kimutatta, hogy “a nagyobb keverési sebesség nagyobb hidrogént, valamint nagyobb sejtsűrűséget eredményezett” az R. rubrum esetében (lásd a 2. ábrát).
Egy másik mikrobiológiai vizsgálatban, amelyet Matthew Ross Melnicki végzett, azt találták, hogy az R. rubrum a hidrogénszintézis révén megújuló energiát, például hidrogén üzemanyagot biztosít. Biológiai útvonalai nagy sebességgel képesek hidrogénszintet előállítani, ha együtt termesztik a Chlamydomonas reinhardtii algafajjal, amellyel az R. rubrum harmonikusan együtt tud nőni. Melnicki a hidrogénszintézisre egy nem hagyományos együttkultivációs megközelítést javasolt, amely a hidrogénszintézis olyan megnövelt szintjeit eredményezte, amelyek nem érhetők el, ha az együttkultivációt nem kapcsolják össze. A C. reinhardtii magasabb fényintenzitás mellett volt előnyös, míg az R. ruburm alacsonyabb fényintenzitás mellett volt termelékenyebb, ami rávilágított a ko-kultiválásuk előnyeire.
A R. rubrum termesztésére egy feed-batch technikát alkalmazva megállapították, hogy a beoltott kultúrák az exponenciális növekedési fázisban hidrogént termeltek, és a hidrogénszintézis 70 órán keresztül folyamatosan virágzott. Amikor a növekedés leállt, Melnicki szukcinátot adott hozzá, amely elektrondonorként működik az anyagcsere-útvonalban, és a hidrogénfejlődés folytatódott, bár a baktérium már nem növekedett tovább. Fontos azonban megjegyezni, hogy a szukcinát folyamatos hozzáadásával csökkent hidrogénfejlődési sebességet tapasztaltak. Amint további növekedési táptalajt adtak hozzá, a hidrogéntermelés sebessége ismét megnőtt.”
Melnicki azt is megállapította, hogy “bár a növekedés nem szükséges a hidrogéntermeléshez, ez a munka a sejtek növekedésének szükségességét igazolja a maximális sebesség fenntartásához, ami a félig folyamatos tenyésztési stratégia ipari alkalmasságára utal” (Melnicki, 2009).
Cool Factor
A Rhodospirillum rubrum genomszekvenciája a Rhodospirillum nemzetségből a R. centenum után csak a második, amelyet közzétettek (Munk et al, 2011).

A Rhodospirillum rubrum érdekes tulajdonsága, hogy különböző körülmények között képes színt változtatni. Ez egy anoxigén fototróf, de nem termel extracelluláris oxigént. Ehelyett elemi ként termel a fénygyűjtés során. Ez annak köszönhető, hogy a R. rubrumból hiányzik a fénygyűjtő 2-es komplex (Munk et al, 2011). A baktérium baktérium-klorofillt használ, ami lehetővé teszi számára, hogy olyan hullámhosszúságú fényt nyeljen el, amelyet a növények nem hasznosítanak. A baktérium-klorofill 800-925 nm-es hullámhosszakat nyel el.
Aerob körülmények között a R. rubrum fotoszintetikus aktivitása genetikailag elnyomódik, és a baktérium színtelenné válik. Anaerob körülmények között azonban a fotoszintézis aktiválódik, és a baktérium határozott lilásvörös színt vesz fel (lásd az 5. képet). Ennek oka a membránban található kromatofórák aktivitása (Munk et al, 2011). A kromatofórák tartalmazzák a baktérium bakterio-klorofill pigmentjeit, és a fotoszintézis aktiválása során abszorpciós változásokon mennek keresztül.
Benson, A. “Chloroplast lipid metabolizmus”. In Symposium no. VI. V. nemzetközi biokémiai kongresszus. 1961. Oxford: Pergamon Press. (In the Press.)
Benson, A., Wintermans, J. és Wiser, R. “Chloroplast lipids as carbohydrate reservoirs”. Plant Physiology. 1959. Volume 34. p. 815.
Bergey, D. and Holt, J. Bergey’s manual of determinative bacteriology. Kilencedik kiadás. Baltimore. Lippincott Williams and Wilkins. 1994. 359. o.
Brock, T.D., M.T. Madigan, J.M. Martinko és J Parker. Biology of Microorganisms. 9. kiadás. Upper Saddle River: Prentice Hall, 2000. p. 459.
Chance, B. and Smith, L. “A Rhodospirillum rubrum légzőszervi pigmentjei”. Nature. Volume 174. p. 803-806.
Cohen-Bazire, G. és Kunisawa, R. “The Fine Structure of Rhodospirillum Rubrum .” Journal of Cell Biology. 1963. Volume 16. p. 401-419.
Hulcher, M. and Conti, S. “Cytochronies in chlorophyll-containing particles of Chromatium and Chlorobium thiosuphatophilum.”. Biokémiai és biofizikai kutatási közlemények. 1960. Volume 3. p. 497.
Kamen, M. “The Haem Proteins of Photosynthetic Tissues”. In Symposium no. VI. V. nemzetközi biokémiai kongresszus. 1961. Oxford: Pergamon Press. (In the Press.)
Keister, D. és Minton, N. “Energy-linked reactions in photosynthetic bacteria. 3. További vizsgálatok az energiához kötött nikotinamid-adenin-dinukleotid redukcióról a Rhodospirillum rubrum kromatofórák által”. Biochemistry. 1969. Volume 8. p. 167-173.
Mastroleo, F., Monsieurs, P., and Leys, N. “Insight into the radiotolerance of the life support bacterium Rhodospirillum rubrum S1H by means of phenotypic and transcriptomic methods”. COSPAR Tudományos Közgyűlés. 2010. 4. o.
Melnicki, M. R. “Modes of hydrogen production in the photosynthetic bacterium, Rhodosphirillum rubrum”. University of California, Berkeley. 2009.
Munk, C., Copeland, A., Lucas, C., Lapidus, A., et al. “Complete genome sequence of Rhodospirillum rubrum type strain (S1T)”. A genomikai tudományok szabványai. 2011. Volume 4. pp. 293-302.
Najafpour, G., Ismail, K., Younesi, H., Mohamed, A. R., and Kamaruddin, A. H. “Hydrogen as clean fuel via continuous fermentation by anaerobic photosynthetic bacteria, Rhodospirillum rubrum”. African Journal of Biotechnology. 2004. Volume 3. pp. 503-507.
Newton, J. és Newton, G. “Composition of the photoactive subcellular particles from Chromatium”. The Archives of Biochemistry and Biophysics. 1957. Volume 71. p. 250.
Oelze, J. és Weaver, P. “The adjustment of photosynthetically grown cells of Rhodospirillum rubrum to aerobic light conditions.” (A Rhodospirillum rubrum fotoszintetikusan növesztett sejtjeinek aerob fényviszonyokhoz való alkalmazkodása). The Archives of Microbiology. 1982. Volume 79. p. 108-121.
Reslewic, S., Shigou, Z., Place, M., Zhang, Y., Briska A., Goldstein, S., Churas, C., Runnheim, R., Forrest, D., Lim, A., Lapidus, A., Han, C., Roberts, G., and Schwartz, D. “Whole-Genome Shotgun Optical Mapping of Rhodospirillum rubrum .” Applied and Environmental Microbiology. 2005. Volume 71. p. 5511-5522.
Schachman, H., Pardee, A., and Stanier, R. “Studies on the macroromolecular organization of microbial cells” (Vizsgálatok a mikrobiális sejtek makromolekuláris szerveződéséről) . The Archives of Biochemistry and Biophysics. 1952. Volume 38. p. 245-260.
Schön, G. és Biedermann, M. “Az illósavak szintézise piruvát és fruktóz fermentációjával Rhodospirillum rubrum anaerob sötét kultúrákban”. Archiv fur Mikrobiologie.1972. Volume 85. p. 77-90.
Schultz, J. és Weaver, P. “Fermentáció és anaerob légzés Rhodospirillum rubrum és Rhodopseudomonas capsulate által”. The Journal of Bacteriology. 1982. Volume 149. p. 181-190.
Ulmer, H., Gross, R., Posada, M., Weisbach, P., Fuller, R. C., and Lenz, R. “Bacterial production of poly(.beta.-hydroxyalkanoates) containing unsaturated repeating units by Rhodospirillum rubrum”. Makromolekulák. 1994. Volume 27. p. 1675-1679.
Weaver, P. “Temperature-Sensitive Mutations of the Photosynthetic Apparatus of Rhodospirillum rubrum”. Proceedings of the National Academy of Sciences of the United States of America. 1971. Volume 68. p. 136-138.
Edited by students of Dr. Iris Keren: Allison Almeida, Matthew Cozzolino, Trung Nguyen, és Jessica Wong.
NEUF2011