- Origins of Triceratops and other Ceratopsid Dinosaurs
- Variation in Frills and Horns of the same species
- The Triceratops Saga… The Great Species Roundup: From 16 species to 2.
- Triceratops vs. Torosaurus – Triceratops ei ole Torosaurus
- Sarvet ja röyhelöt – Niitä ei tehty ramppaamiseen!
- Triceratopsin käyttäytyminen: Herding and Diet
- Herding Behavior
- Hampaat ja ruokavalio
- Kuinka nopeasti Triceratops-dinosaurus pystyi juoksemaan?Triceratopsin ja keratopsidien liikkuminen
- Triceratops-dinosauruksen iho – suomuja, suomuja ja sulkakynsiä??
- Suositeltavat Tyrannosauruskirjat ja -fossiilit:
Origins of Triceratops and other Ceratopsid Dinosaurs

Protoceratops andrewsi specimen at the CMNH. Tämä pieni, kaksimetrinen keratopsidi on yksi harvoista aasialaisista keratopsidi-dinosauruksista.
Keratopsidit – Alkuperä ja levinneisyys
Keratopsidae-dinosaurusten suku käsittää nelijalkaiset kasvinsyöjät, joilla on tunnusomaiset kasvojen sarvet ja suuret röyhelöt. Tähän dinosaurusperheeseen kuuluu tunnettu Triceratops, joka on ensimmäinen vuonna 1887 löydetty Ceratopsid. Ceratopsideihin kuuluu lukuisia alahahmoja ja sukuja. Ne luokitellaan sarvien, nokkien ja röyhelöiden muodon ja koon perusteella.
Ceratopsidit rajoittuvat Pohjois-Amerikkaan ja Aasiaan (Triceratopsia tavataan vain läntisessä Pohjois-Amerikassa). Alkukantaisia keratopsideja on löydetty Aasiasta, mikä viittaa siihen, että ne ilmestyivät ensin sinne ja siirtyivät sitten maasillalla Pohjois-Amerikkaan. Pohjois-Amerikassa ne monipuolistuivat nopeasti, kunnes ne kuolivat sukupuuttoon liitukauden lopulla. Ceratopsideja on luokiteltu noin 37 sukua. Alla on kuva muutamien Ceratopsid-sukujen kalloista, joista näkyy selvästi erilaiset sarvien ja hapsujen muodot ja koot.

Kuva eräiden Ceratopsid-dinosaurusten suvun jäsenten kalloista. Huomaa hapsujen, nokan ja sarvien hurja vaihtelu. Kuva on peräisin Utahin luonnonhistoriallisesta museosta.
By skinnylawyer from Los Angeles, California, USA , via Wikimedia Commons
Variation in Frills and Horns of the same species
Kun Ceratopsideja löydettiin 1900-luvun alkupuolella, sirpaleisia jäännöksiä luokiteltiin niiden hapsu-, nokka- ja sarvimuotojen perusteella. Nuorten ja aikuisten ulkonäköä ja sukupuolista dimorfismia ei otettu huomioon. Tämän seurauksena pystytettiin lukemattomia lajeja.
Myöhemmin, kun fossiilisia todisteita löydettiin yhä enemmän, paleontologit tajusivat, että kallon muoto muuttuu eläimen siirtyessä eri kehitysvaiheisiin. Sarvet ja röyhelöt kehittyivät täysin vasta aikuisiällä (Farlow 327).
Hyvä esimerkki tästä on vuonna 2013 löydetystä vauvasta chasmosaurine ceratopsid dinosauruksesta. Currie et al. julkaisivat vuonna 2016 artikkelin tästä vauvadinosauruksesta ja vertasivat sitä aikuisiin chasmosarusnäytteisiin (Currie et al., 2016). Vauva-chasmosarusin frill näyttää täysin erilaiselta kuin aikuisen frill.
The Triceratops Saga… The Great Species Roundup: From 16 species to 2.

A Triceratops Dinosaur skeleton on esillä Field Museum of Natural History -museossa Chicagossa
Hyvä esimerkki Ceratopsidien määrän vähentämisestä ovat Triceratops-suvut. Triceratops on yleisimmin löydetty dinosaurus Yhdysvaltojen länsiosien Hell Creek -muodostumassa. Triceratops-sukuun luokiteltiin aikoinaan jopa 16 lajia kaikkien löydettyjen erilaisten kallovariaatioiden perusteella. Kun paleontologit huomasivat, että röyhelöt muuttuivat kehityksen aikana, Triceratops analysoitiin uudelleen. Vuonna 1986 Ostrom ja Wellnhofer supistivat Triceratopsin vain yhteen lajiin, T. horridus. He päättelivät, että kaikki kallojen erot johtuivat eri kehitysvaiheista ja sukupuolisesta dimorfismista, eikä samalla alueella voinut elää lukuisia samankaltaisia suuria eläimiä.
Hieman myöhemmin Catherine Forester vuonna 1996 tutki Triceratopsin lajeja ja päätyi siihen, että koska T. horridus oli huomattavasti suurempi kuin T. prorsus, ne olivat kaksi eri lajia. Lopulta vuonna 2009 Scannella ja Fowler tukivat tätä näkemystä, ja saaga oli ohi. Triceratops-lajeja oli kaksi… tavallaan… Sitten on vielä tuo Torosaurus-kysymys…
Triceratops vs. Torosaurus – Triceratops ei ole Torosaurus

Triceratops vs. Torosaurus – Triceratops ei ole Torosaurus
Torosaurus latus -näytekappaleiden kokovertailu.
”A, YPM 1831, subadultti, kallon pituus >2,6 m, ja B, ANSP 15192, varhainen aikuinen, kallon pituus 1,8 m. YPM 1831:n rostrum on rekonstruoitu ja se olisi todennäköisesti ollut hieman pidempi, kuten ANSP 15192:lla.” From: Nicholas R. Longrich*, Daniel J. Field, 2012 – PLOS one CC2-lisenssillä.
Torosaurus on suuri keratopsidi, joka on löydetty samasta muodostumasta kuin Triceratops. Se on kookkaampi, ja sen frill on selvästi erilainen, pidempi ja siinä on suuria reikiä lähellä yläosaa. Vuonna 2010 Scannella ja Horner julkaisivat artikkelin, jossa osoitettiin, että Torosaurus oli itse asiassa Triceratopsin myöhäinen, nopea kasvuvaihe. Tämä herätti monenlaisia kiistoja. Paleontologit ottivat kantaa, jotkut olivat samaa mieltä ja jotkut eri mieltä. Henkilökohtaisesti minulla on aina ollut vaikeuksia sen kanssa, miten kaksi suurta reikää voisi yksinkertaisesti syöpyä hapsiluuhun. Se todella muuttaisi koko lihasten kiinnitysrakenteet. Viime vuosina on julkaistu monia artikkeleita, jotka kumoavat Scannellan ja Hornerin tulokset. Vuonna 2011 keratopsidi-dinosaurusten asiantuntija tohtori Andy Farke julkaisi artikkelin, joka osoitti Torosauruksen olevan ”Triceratopsista ja Nedoceratopsista erillinen suku”. Viimeaikaiset artikkelit, kuten Longrichin ja Fieldin vuonna 2012 julkaisema artikkeli, tukevat pikemminkin erillistä sukua. He tutkivat erilaisia Torosaurus- ja Triceratops-kalloja ja tekivät klusterointianalyysin järjestääkseen ne kasvusarjoihin. He löysivät sekä Torosauruksen että Triceratopsin epäkypsiä ja täysikasvuisia yksilöitä. Siksi ne saattoivat olla sama eläin. Triceratops ja Torosaurus ovat eri dinosauruksia.
Yllä oleva kuva on Longrichin ja Fieldin (2012) teoksesta. Siinä näkyy aikuinen ja subadultti Torosoarus.
Sarvet ja röyhelöt – Niitä ei tehty ramppaamiseen!

Ceratopsidi-dinosauruksen kallo Utahista, jossa näkyy sen taidokkaat sarvet ja röyhelöt CMNH:n näytteillä
Pääosalla Ceratopsideista oli valtavat röyhelöt lukuun ottamatta Diceratopsia ja Triceratopsia. Näillä kahdella suvulla oli verrattain pienikokoiset frillit (Fastovsky s. 180). Hapsujen koon kasvu elämän aikana vaikutti suoraan leukalihasten voimaan (mitä suurempi hapsu, sitä suurempi/vahvempi leukalihas) (Fastovsky s. 186). Joissakin hapsuissa nähdyt suuret reiät oli luultavasti täytetty lihasten kiinnikkeillä ja/tai rustolla.
Sarvien osalta monet ihmiset unohtavat nähdessään fossiilisen Triceratopsin tai muun keratopsidi-dinosauruksen, että kallossa olevat sarvet ovat luun ydintä. Nämä luunsydämet olisivat olleet varsinaisten sarvien peitossa, jotka olisivat olleet elämässä paljon pidemmät.
Ei valitettavasti ole mahdollista todentaa, mihin sarvia ja röyhelöitä todella käytettiin. Jos kuitenkin käytetään nykyaikaisia paimentolaiseläimiä vertailukohtana, on todennäköistä, että niitä käytettiin dominointiin, puolustautumiseen ja näyttämiseen. Keratopsidit ovat saattaneet käyttää sarviaan oman lajinsa sisällä reviiri- ja pariutumisoikeuksiin. Tämä sopii yhteen fossiilisten todisteiden kanssa, sillä Triceratopsin kalloista ja hapsuista on yleisesti löydetty pistohaavoja toisilta Triceratopsilta, mikä viittaa jonkinlaiseen lajinsisäiseen kamppailuun (Farke, et al., 2009).
Tutkimus osoitti, että jos kaksi aikuista Triceratopsia törmäisi toisiaan päin, niiden kallot hajoaisivat! Ceratopsidien sarvet olivat kuitenkin muodoltaan erilaiset kuin nisäkkäiden vastaavat. Ryntäämisen sijaan ne saattoivat yksinkertaisesti lukita sarvet yhteen ja taistella. Mallien avulla tehdyssä tutkimuksessa todettiin, että triceratopsin oli fyysisesti mahdollista käyttää sarvien lukitusta (Farke, Andrew, A. 2004). He löysivät 3 mahdollista sarvien lukitusjärjestelyä. Sen sijaan, että ne olisivat rampanneet toisiaan, ne lukitsivat sarvet ja tappelivat, vähän samaan tapaan kuin hirvet lukitsevat sarvet taistellessaan.
On mielenkiintoista huomata, että on olemassa yksi poikkeus. Farke (Farke, et al., 2009) tutki erästä toista sukua, Centrosaurusta, ceratopsidia, jolla oli vain yksi suuri nenäsarvi, ja sen todettiin olevan vailla vammoja. Tämä tarkoittaa, että Centrosaurukset eivät todennäköisesti käyttäneet sarviaan ja röyhelöitään paritteluun tai reviiritaisteluun, vaan sen sijaan näyttötarkoituksiin, kuten riikinkukko. Alla on kuvattu kuva Farken artikkelista. Siinä on esitetty vaurioiden esiintyvyysasteet (A) Triceratopsissa ja (B) Centrosauruksessa.
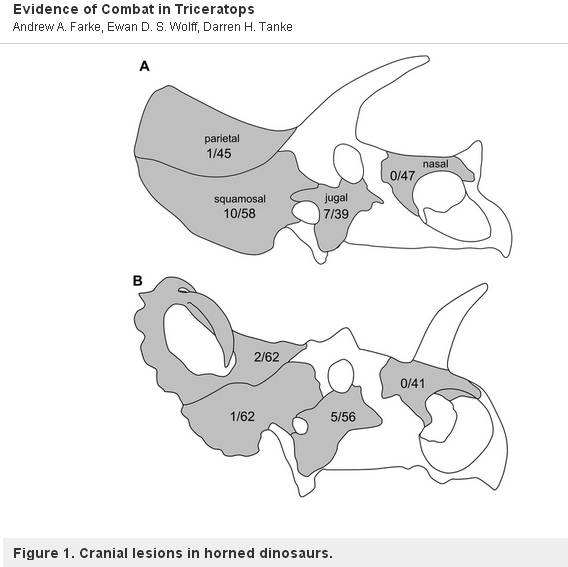
”Schematics of the skulls of (A) Triceratops and (B) Centrosaurus, showing incidence rates of lesions (periosteal reactive bone and fracture calluses) on each cranial element and (epänormaalien elementtien määrä / elementtien kokonaismäärä).”. Ei mittakaavassa.”
From: Andrew A. Farke, Ewan D. S. Wolff, Darren H. Tanke, 2009
– PLOS one CC2-lisenssillä.
Tarinan moraali, jos Triceratops käytti niitä pariutumiskäyttäytymiseen ja Centrosaurus näytöskäyttäytymiseen, sarvia ja röyhelöitä käytettiin luultavasti useilla eri ceratopsidatyypeillä monin eri tavoin.
Yksi monien ceratopsidien sarvien ja röyhelöiden käyttötapa oli luultavasti saalistajien karkottaminen. Joistakin ceratopsidi-fossiileista on löydetty jopa parantuneita T-rexin puremajälkiä, nämä ovat yksilöitä, jotka ovat onnistuneesti torjuneet T-rexin hyökkäyksen. Monissa toisissa taas on löydetty parantumattomia puremajälkiä, mikä tarkoittaa, että ne olivat T-Rexin ravintoa. Näiden fossiililöytöjen perusteella ei ole mahdoton olettaa, että hapsuja ja sarvia käytettiin myös torjumaan mahtavaa T-Rexiä, joka asui Yhdysvaltojen länsiosissa liitukauden loppupuolella.
Käytettiinpä sarvia ja hapsuja sitten sarvien lukitsemiseen, päähän kiinnittämiseen, esittelyyn, puolustautumiseen tai kaikkeen edellä mainittuun, yksi asia on varma. Niitä oli kaikenmuotoisia ja -kokoisia, katsokaa vaikka alla olevaa kuvaa!

Ceratopsid Dinosauruksen sarvet ja röyhelöt esillä Carnegie Museum of Natural History -museossa.Huomaa, että ensimmäisellä kallolla vasemmalla (Pachyrhinosaurus sp.) ei ole lainkaan sarvea, vaan sillä on sen sijaan paksu luunuppi.
Triceratopsin käyttäytyminen: Herding and Diet
Herding Behavior
Keskilännestä on löydetty suuria luukerrostumia, joissa on satoja Centrosaur-fossiileja, mikä viittaa siihen, että ne kulkivat suurissa laumoissa. Triceratopsista ei ole löydetty suuria luukerrostumia, mutta yksilöiden ryhmiä on löydetty yhdessä, joten ne ovat saattaneet liikkua suurissa laumoissa tai ainakin pienemmissä ryhmissä. tämä on sopusoinnussa sen tosiasian kanssa, että sarvien käyttö on yleisempää paimentavilla eläimillä, kuten peuroilla, gaselleilla ja puhvelilla.

Kuva kolmesta Triceratopsin hampaasta, jotka löydettiin Etelä-Dakotan Hell Creek -muodostumasta Hell Creek -dinosauruskaivaukseni aikana.
Hampaat ja ruokavalio
Kuten kaikki paimentolaiseläimet, Ceratopsidit olivat kasvinsyöjiä. Niiden suu soveltui erinomaisesti sitkeän kasvillisuuden syömiseen. Näillä dinosauruksilla oli kaksoisjuurihampaat, jotka olivat järjestäytyneet ryhmiin, joita kutsutaan hammaspattereiksi. Hammaspatteri on ainutlaatuinen hammasasetelma, jossa vierekkäiset hampaat olivat lukittuneet toisiinsa pituussuuntaisiksi riveiksi ja pystysuoriksi sarakkeiksi. Jokaisessa pylväässä oli vähintään 3 hammasta, joista 1 oli toiminnallinen ja 2 oli ”korvaavia”. Hampaat, joita vaihdettiin jatkuvasti koko dinosaurusten elämän ajan, oli sijoitettu niin pystysuoraan ja ne olivat niin teräviä (itse asiassa itsestään teroittuvia), että ne pystyivät leikkaamaan kuin sakset. Pureskelun aikana näiden dinosaurusten hampaat eivät todellakaan koskettaneet toisiaan lainkaan, kuten ihmisten hampaat, vaan ne liikkuivat toistensa ohi viiltävällä liikkeellä (Fastovsky 183). Näiden partaveitsenterävien hampaiden ja erittäin vahvojen leukalihasten vuoksi niitä kutsutaan usein ”Luonnon ensimmäisiksi keittiömestareiksi!”. (Farlow 85). Ne pystyivät leikkaamaan läpi kovimmankin materiaalin.
Sivuhuomautuksena mainittakoon, että toisilla yleisillä kasvinsyöjillä, hadrosauruksilla, oli samanlaiset hammaspatterit.
Tutkijat ovat jokseenkin epävarmoja niiden tarkasta kasvisruokavaliosta. He epäilevät, että suhteellisen matalan haukkumiskorkeutensa vuoksi ne saattoivat syödä matalakasvuisia kasveja, kuten angiospermejä, saniaisia ja pieniä havupuita, jotka kasvoivat sillä korkeudella. Suurten ja painavien kallojensa vuoksi ne eivät voineet nousta takajaloilleen, mutta nämä Amerikan lännen ruohonleikkurit saattoivat pystyä puskemaan puiden yli päästäkseen käsiksi lehtiin.

Kuva Triceratopsin hampaasta, joka on edelleen upotettu Hell Creek -muodostumaan Hell Creek -dinosauruskaivaukseni aikana.
Kuinka nopeasti Triceratops-dinosaurus pystyi juoksemaan?Triceratopsin ja keratopsidien liikkuminen
Monet olettavat, että Triceratops oli hyvin hidas ja kömpelö eläin. Todisteet osoittavat, että näin ei ole.
Monissa museoissa olevat Ceratopsid-dinosaurusten luurankorekonstruktiot (mukaan lukien tässä artikkelissa kuvatut) esittävät Triceratopsin rönsyilevän liskon kaltaisessa asennossa.
Tutkimukset osoittavat, että näillä dinosauruksilla olisi pitänyt olla nisäkkäiden kaltaisempi asento, jossa etujalkojen olisi pitänyt olla eläimen alapuolella, ei levittäytyneinä sivulle. Liskon kaltainen asento johtuu virheistä Ceratopsidien kylkiluiden ja nikamien kiinnityksessä.
Todisteet, jotka tukevat edellä esitettyjä väitteitä, ovat peräisin Triceratopsin ja muiden Ceratopsidien jälkien analysoinnista. Paul ja Christiansen julkaisivat vuonna 2000 artikkelin, jossa he tutkivat jälkien jalkajälkiä jälkijäljissä ja analysoivat käsien ja jalkojen sijoittelua ja kulmia. Näiden tietojen perusteella he päättelivät, että Ceratopsideilla on norsun kaltainen eturaajojen asento. Nisäkkään kaltainen asento antaa Ceratopsideille mahdollisuuden juosta PALJON nopeammin kuin matelijoiden asento. Paul ja Christiansen ilmoittavat sitten suurimpien Ceratopsidien maksimijuoksunopeuden olevan ”suurin piirtein samanlainen kuin sarvikuonoilla”. Musta sarvikuono voi muuten juosta noin 55 kilometrin tuntinopeudella eli 34 mailia tunnissa! Se on aika pirun nopea!

Carnegie Museum of Natural History -museon Triceratopsin luurankorekonstruktio osoittaa rönsyilevää, liskon kaltaista ryhtiä, mikä viimeaikaisten todisteiden mukaan ei pidä paikkaansa.

Chicagossa sijaitsevan Field-museon Triceratopsin pienoismalli osoittaa pikemminkin nisäkkäiden kaltaista ryhtiä, mikä viimeaikaisten todisteiden mukaan pitää paikkansa.
Triceratops-dinosauruksen iho – suomuja, suomuja ja sulkakynsiä??
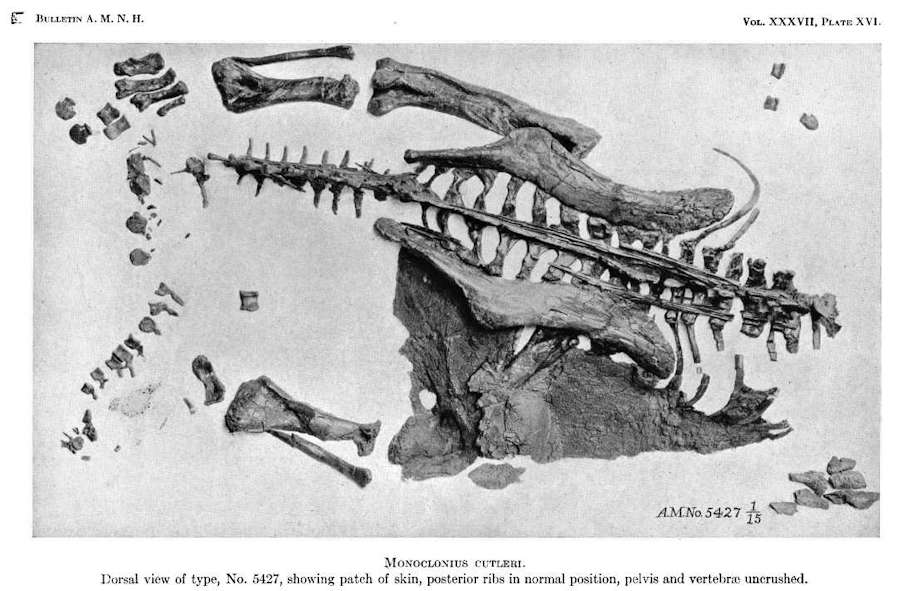
Kuva Centrosaurus-dinosauruksen näytteestä (AMNH 5427) täydellisenä, jossa on suuri nahkaläiskä
Ajankohta: 1917, Tekijä: Barnum Brown,Lähde: Art: AMNDH Digital Library: B037a10.pdf Public Domain.
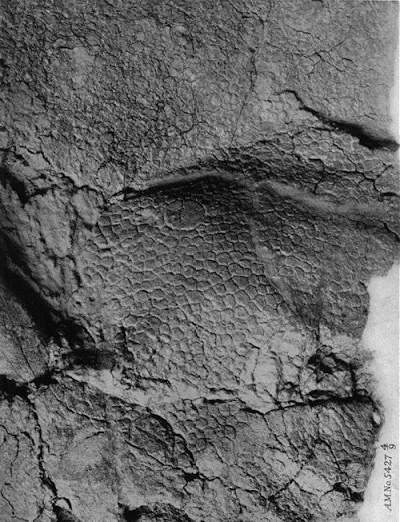
Lähikuva Centrosaurus-dinosauruksen (AMNH 5427) nahanjäljestä (Centrosaurus dinsoauruksen nahanjälki).
Ajankohta: 1917, Tekijä: Barnum Brown,Lähde: KUVAILULEHTI 1: B037a10.pdf Public Domain.
Fossiilisoituneita dinosauruksen ihonjälkiä on erittäin harvoin löydettävissä. Iholla varustettuja dinosaurusfossiileja tituleerataan usein ”dinomuumioiksi”, vaikka ne eivät ole muumioituneita. Varsinainen iho ei ole säilynyt, mutta eläintä aikoinaan ympäröineeseen pehmeään mutaan jääneet ihojäljet fossiilisoituvat.
Triceratopsin ystäville onneksi on löydetty muutamia ”Ceratopsid-muumioita”. Yksi alkuperäisistä, yllä kuvassa AMNH 5427, on Centrosaurus, jossa on säilynyt suuri pala ihoa.
Uudempi ”Triceratops-muumio” nimeltä Lane löydettiin Wyomingista vuonna 2002. Black Hills Institute of Geological Research kaivoi T. horridus -näytteen huolellisesti ja vaivalloisesti esiin. Se on nyt esillä heidän museossaan yhdessä toisen lähes täysin ehjän T. horriduksen luurangon, Kelseyn, kanssa.
Triceratopsin kaistalla on parhaiten säilynyt iho kuin millään muulla löydetyllä dinosaurusfossiililla. On alueita, joissa on säilynyt useita metrejä ihoa.
Nahka on hyvin erilainen kuin mitä monet paleontologit luulivat. Eläimen alapuolta peittävät krokotiilin kaltaiset suomut. Muu osa dinosauruksesta on pienten suomujen peitossa. Siellä on kuitenkin myös epäsäännöllisiä suomujen kaltaisia muotoja. Jotkut ovat nyrkinmuotoisia kuusikulmioita, ja niiden keskellä on pieniä huokosmaisia reikiä.
Jotkut ovat arvelleet, että näistä rei’istä olisi voinut työntyä ulos sulkakynsiä. Tämä ”sulkakynsi”-hypoteesi on peräisin Triceratopsin sukulaiselta Psittacosaurukselta Aasiasta. Se on kaukainen sukulainen, jonka pyrstön yläosassa on sulkakynän kaltaisia rakenteita. On tärkeää huomata, että Psittacosaurur on Triceratopsin kaukainen sukulainen, joten se ei tarkoita, että myös Triceratopsilla olisi ollut sulkakynät. ”Quill”-hypoteesi on tällä hetkellä spekulaatioiden aluetta, ja meidän on odotettava lisätutkimuksia ja julkaisuja, jotka valaisevat Triceratopsin ihon tarkkaa luonnetta. Alla on kuva nahkaleikkauksesta th Triceratops Laneista.

Section of the Triceratops skin cast from the dinosaur Lane showing the hexagon scales. Kuva: Black Hills Institute of Geological Research

Psittacosaurus-dinosaurus säilyneiden sulkakynien ja mallin kanssa. Tämä on Triceratopsin kaukainen sukulainen.paleontologit arvelevat, että sulkarakenteet ovat voineet työntyä esiin joistakin Triceratopsin oudoista suomuista.
Suositeltavat Tyrannosauruskirjat ja -fossiilit:

New Perspectives on Horned Dinosaurs: The Royal Tyrrell Museum Ceratopsian Symposium (Life of the Past)
Tässä kirjassa annetaan päivitetty näkemys ceratopsioista. Siinä esitellään uutta tutkimusta sarvipäisten dinosaurusten toiminnasta, biologiasta, käyttäytymisestä, paleoekologiasta ja paleogeografiasta. Siinä kuvataan myös monia uusia lajeja. Kirja on kokoelma tieteellisiä artikkeleita, joten tämä on tarkoitettu vain vakavasti otettaville dinosaurusten lukijoille! Mielestäni se on loistava hakuteos ja pakollinen tietolähde dinosaurusten harrastajille.

Dinosaurukset uudelleen löydetty: The Scientific Revolution in Paleontology
by Michael J. Benton, 2019
Tämä on hyvä yleiskirja dinosauruksista. Se on ajan tasalla ja käy läpi viimeisen 20 vuoden aikana tehdyt uudet löydöt, jotka ovat muuttaneet täysin näkemyksemme dinosauruksista. Kirja on jaettu helppoihin lukuihin, joissa käydään läpi niiden värejä, nopeuksia, puremisvoimaa, lastenhoitoa ja paljon muuta… asioita, joita emme tienneet vielä vuosia sitten!Tutustu arvosteluihin ja näytesivuihin.

Laadukkaat dinosauruksen hampaat by Fossilera
Viittaukset / siteeratut teokset
Currie Philip J., Holmes Robert B., J. Ryan Michael, Coy Clive. (2016) A juvenile chasmosaurine ceratopsid (Dinosauria, Ornithischia) from the Dinosaur Park Formation, Alberta, Canada. Journal of Vertebrate Paleontology, 2016; e1048348 DOI: 10.1080/02724634.2015.1048348
Farlow, O. James, Brett-Surman, M.K., Editors (1997). The Complete Dinosaur. IN: Indiana University Press.
Fastovsky, E. David, Weishampel, B. David (1996). The Evolution And Extinction Of The Dinosaurs. NY: Cambridge University Press.
Farke A.A. (2011) Anatomy and Taxonomic Status of the Chasmosaurine Ceratopsid Nedoceratops hatcheri from the Upper Cretaceous Lance Formation of Wyoming, U.S.A. PLoS ONE 6(1): e16196. doi: 10.1371/journal.pone.0016196
Farke AA, Wolff EDS, Tanke DH. (2009) Evidence of Combat in Triceratops. PLoS ONE 4(1): e4252. doi: 10.1371/journal.pone.0004252
Farke, A.A. 2004. Horn use in Triceratops (Dinosauria: Ceratopsidae): Testing behavioral hypotheses using scale models. Palaeontologia Electronica 7(1):10 pp. (PDF)
Forster Catherine A. (1996) Species resolution in Triceratops: cladistic and morphometric approaches. Journal of Vertebrate Paleontology, 16:259-270.
Longrich NR, Field DJ. (2012) Torosaurus ei ole Triceratops: Ontogeny in Chasmosaurine Ceratopsids as a Case Study in Dinosaur Taxonomy. PLoS ONE 7(2): e32623. doi:10.1371/journal.pone.0032623
Ostrom J.H, Wellnhofer P. (1986) The Munich specimen of Triceratops with a revision of the genus. Zitteliana. 14, 111-158.
Scannella J, Horner JR. (2010) Torosaurus on Triceratops, synonymy through ontogeny. Journal of Vertebrate Paleontology 30: 1157-1168.
Scannella, J.B. ja D.W. Fowler. (2009) Anagenesis in Triceratops: evidence from a newly resolved stratigraphic framework for the Hell Creek Formation. North American Paleontological Convention , abstracts volume: 148-9.
Gregory S. Paul, Per Christiansen. (2000) Forelimb posture in neoceratopsian dinosaurs: implications for gait and locomotion Paleobiology Summer, v. 26 no. 3 p. 450-465