Typpeä sisältävät emäkset, nukleotidien tärkeät komponentit, ovat orgaanisia molekyylejä, ja ne ovat saaneet nimensä hiilestä ja typestä. Ne ovat emäksiä, koska ne sisältävät aminoryhmän, joka voi sitoa ylimääräisen vedyn ja siten vähentää vetyionien pitoisuutta ympäristössään tehden siitä emäksisemmän. Jokainen DNA:n nukleotidi sisältää yhden neljästä mahdollisesta typpiperäisestä emäksestä: adeniini (A), guaniini (G), sytosiini (C) ja tymiini (T). Myös RNA:n nukleotidit sisältävät yhden neljästä mahdollisesta emäksestä: adeniini, guaniini, sytosiini ja urasiili (U) tymiinin sijasta.
Adeniini ja guaniini luokitellaan puriineiksi. Puriinin perusrakenne on kaksi hiili-typpirengasta. Sytosiini, tymiini ja urasiili luokitellaan pyrimidiiniksi, joiden primaarirakenteena on yksi hiili-typpirengas (kuva 1). Kuhunkin näistä hiili-typpirenkaista on kiinnittynyt erilaisia funktionaalisia ryhmiä. Molekyylibiologian lyhenteessä typpiperustat tunnetaan yksinkertaisesti symboleilla A, T, G, C ja U. DNA:ssa on A, T, G ja C, kun taas RNA:ssa on A, U, G ja C.
DNA:ssa pentoosisokeri on deoksiriboosi, ja RNA:ssa sokeri on riboosi (kuva 1). Sokereiden erona on hydroksyyliryhmän läsnäolo riboosin toisessa hiilessä ja vedyn läsnäolo deoksiriboosin toisessa hiilessä. Sokerimolekyylin hiiliatomit on numeroitu seuraavasti: 1′, 2′, 3′, 4′ ja 5′ (1′ lukee ”yksi prime”). Fosfaattijäämä on kiinnittynyt yhden sokerin 5′-hiilen hydroksyyliryhmään ja seuraavan nukleotidin sokerin 3′-hiilen hydroksyyliryhmään, mikä muodostaa 5′-3′-fosfodiesterisidoksen. Fosfodiesterisidos ei muodostu yksinkertaisella dehydraatioreaktiolla kuten muut makromolekyylien monomeerejä yhdistävät sidokset: sen muodostuminen edellyttää kahden fosfaattiryhmän poistamista. Polynukleotidissa voi olla tuhansia tällaisia fosfodiesterisidoksia.
DNA:n kaksois-Helix-rakenne
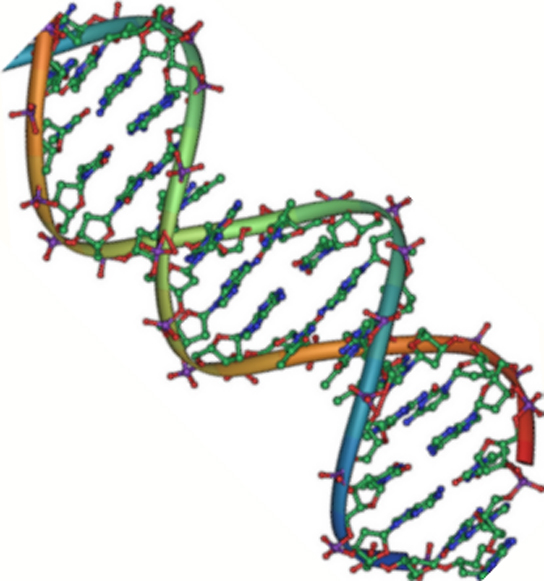
Kuva 2. DNA on antiparalleelinen kaksoiskierre. Fosfaattirunko (kaarevat viivat) on ulkopuolella, ja emäkset ovat sisäpuolella. Kukin emäs on vuorovaikutuksessa vastakkaisen säikeen emäksen kanssa. (luotto: Jerome Walker/Dennis Myts)
DNA:lla on kaksoiskierteinen rakenne (kuva 2). Sokeri ja fosfaatti sijaitsevat kierteen ulkopuolella ja muodostavat DNA:n selkärangan. Typpiperäiset emäkset ovat pinoutuneet sisäpuolelle kuin portaiden askelmat pareittain; parit ovat sitoutuneet toisiinsa vetysidoksilla. Kaksoiskierteen jokainen emäspari on 0,34 nm:n etäisyydellä seuraavasta emäsparista.
Kierteen kaksi säiettä kulkevat vastakkaisiin suuntiin, mikä tarkoittaa, että toisen säikeen 5′-hiilipää on vastakkaisen säikeen 3′-hiilipäähän päin. (Tätä kutsutaan antiparalleeliseksi orientaatioksi, ja se on tärkeää DNA:n replikaatiossa ja monissa nukleiinihappojen vuorovaikutuksissa.)
Vain tietyntyyppiset emäsparit ovat sallittuja. Esimerkiksi tietty puriini voi muodostaa parin vain tietyn pyrimidiinin kanssa. Tämä tarkoittaa, että A voi muodostaa parin T:n kanssa ja G voi muodostaa parin C:n kanssa, kuten kuvassa 3 on esitetty. Tämä tunnetaan nimellä emäskomplementtisääntö. Toisin sanoen DNA-juosteet ovat toisiaan täydentäviä. Jos yhden säikeen sekvenssi on AATTGGCC, komplementaarisen säikeen sekvenssi on TTAACCGG. DNA:n replikaation aikana kumpikin säie kopioituu, jolloin syntyy tytär-DNA:n kaksoiskierre, joka sisältää yhden vanhemman DNA-juosteen ja uuden syntetisoidun juosteen.
Harjoituskysymys
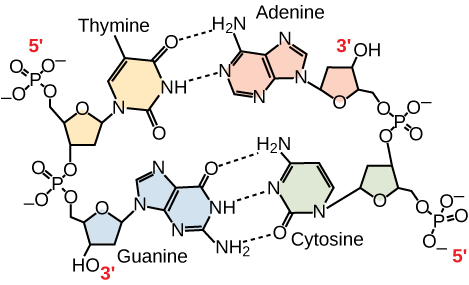
Kuvio 3. DNA:n replikaatio. Kaksisäikeisessä DNA-molekyylissä kaksi säiettä kulkevat antiparalleelisesti toisiinsa nähden siten, että toinen säie kulkee 5′-3′ ja toinen 3′-5′. Fosfaattirunko sijaitsee ulkopuolella, ja emäkset ovat keskellä. Adeniini muodostaa vetysidoksia (eli emäspareja) tymiinin kanssa, ja guaniini muodostaa emäspareja sytosiinin kanssa.
Mutaatio tapahtuu, ja sytosiini korvataan adeniinilla. Minkä vaikutuksen arvelet tällä olevan DNA:n rakenteeseen?
RNA
Ribonukleiinihappo eli RNA osallistuu pääasiassa proteiinisynteesiin DNA:n johdolla. RNA on yleensä yksijuosteista ja koostuu ribonukleotideista, jotka ovat yhteydessä toisiinsa fosfodiesterisidoksilla. RNA-ketjun ribonukleotidi sisältää riboosia (pentoosisokeria), yhden neljästä typpiperäisestä emäksestä (A, U, G ja C) sekä fosfaattiryhmän.
RNA:ta on neljää päätyyppiä: sanansaattaja-RNA (mRNA, messenger RNA), ribosomaalinen RNA (rRNA, ribosomalinen RNA), siirto-RNA (transfer RNA, transfer RNA, tRNA) ja mikroRNA (miRNA, microRNA). Ensimmäinen, mRNA, kuljettaa DNA:n viestiä, joka ohjaa kaikkia solun toimintoja solussa. Jos solu vaatii tietyn proteiinin syntetisointia, tämän tuotteen geeni ”kytkeytyy päälle” ja sanansaattaja-RNA syntetisoidaan ytimessä. RNA:n emäsjärjestys on komplementaarinen sen DNA:n koodausjärjestyksen kanssa, josta se on kopioitu. RNA:ssa ei kuitenkaan ole emästä T, vaan sen tilalla on U. Jos DNA-juosteessa on sekvenssi AATTGCGC, komplementaarisen RNA:n sekvenssi on UUAACGCG. Sytoplasmassa mRNA on vuorovaikutuksessa ribosomien ja muun solukoneiston kanssa (kuva 4).
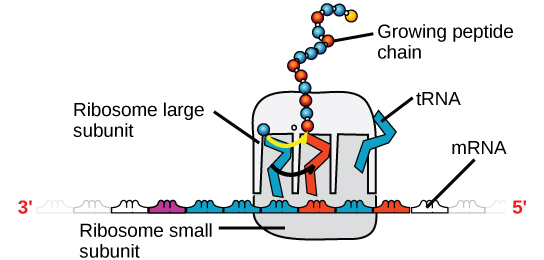
Kuva 4. MRNA:n ja ribosomien välinen vuorovaikutus. Ribosomissa on kaksi osaa: suuri alayksikkö ja pieni alayksikkö. MRNA istuu näiden kahden alayksikön välissä. TRNA-molekyyli tunnistaa mRNA:ssa olevan koodonin, sitoutuu siihen komplementaarisen emäsparin avulla ja lisää oikean aminohapon kasvavaan peptidiketjuun.
MRNA luetaan kolmen emäksen sarjoina, joita kutsutaan koodoneiksi. Kukin kodoni koodaa yhtä aminohappoa. Näin mRNA luetaan ja proteiinituote valmistetaan. Ribosomaalinen RNA (rRNA) on tärkeä osa ribosomeja, joihin mRNA sitoutuu. RRNA varmistaa mRNA:n ja ribosomien oikean linjauksen; ribosomin rRNA:lla on myös entsymaattinen aktiivisuus (peptidyylitransferaasi) ja se katalysoi peptidisidosten muodostumista kahden linjautuneen aminohapon välillä. Transfer-RNA (tRNA) on yksi pienimmistä neljästä RNA-tyypistä, yleensä 70-90 nukleotidin pituinen. Se kuljettaa oikean aminohapon proteiinisynteesipaikalle. Juuri tRNA:n ja mRNA:n välinen emäspariutuminen mahdollistaa oikean aminohapon asettumisen polypeptidiketjuun. mikroRNA:t ovat pienimpiä RNA-molekyylejä, ja niiden tehtävänä on geeniekspression säätely häiritsemällä tiettyjen mRNA-viestien ilmentymistä.