Pagină din Biorealul microbian despre genul Rhodospirillum rubrum
Clasificare
Taxoni de ordin superior
Regat: Bacteria
Filum: Proteobacteria
Clasa: Alphaproteobacteria
Ordine: Alphaproteobacteria: Rhodospirillales
Familia: Rhodospirillaceae
Genul: Rhodospirillum
Specie
Rhodospirillum rubrum
Descriere și semnificație
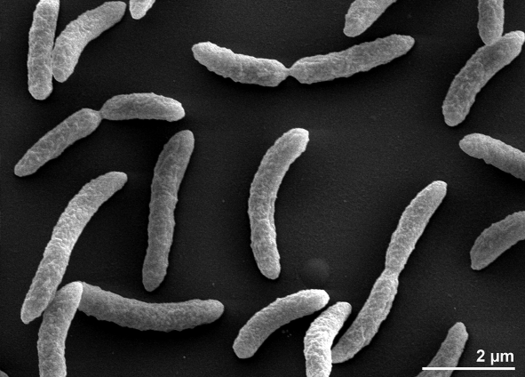
Rhodospirillum rubrum este o proteobacterie Gram-negativă, mezofilă. Temperatura sa optimă de creștere este de 25-30 grade Celsius. Are învelișuri exterioare cu mai multe straturi, care conțin în mare parte grăsimi nesaturate, dar și unele saturate în peretele său celular. R. rubrum este o spirilă, ceea ce înseamnă că are o formă spiralată. Este flagelat polar și, prin urmare, este mobil. Lungimea sa este de 3-10 um, cu o lățime de 0,8-1,0 um.
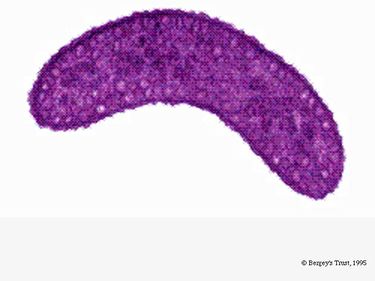
R. rubrum este un anaerob facultativ. În funcție de prezența oxigenului, poate suferi fermentație alcoolică sau respirație aerobă. De asemenea, este capabil de fotosinteză și conține carotenoide și baterioclorofilă în particulele sale cromatofore. Aceste molecule ajută la absorbția luminii și la transformarea ei în energie și, de asemenea, îi conferă culoarea distinctă roșu-violet în condiții anaerobe. R. rubrum este incolor în condiții aerobe.
Deși fotosinteza este activă în condiții aerobe, ea este în general suprimată în prezența O2. Sulful este un subprodus major al fotosintezei, nu O2. R. rubrum poate crește heterotrofic sau autotrofic atunci când face fotosinteză. Spre deosebire de multe plante, R. rubrum nu conține clorofilă a (spectre de absorbție 430-662 nm). Cu toate acestea, conține clorofilă b (spectre de absorbție 660-680 nm) și bacterioclorofile (800-925 nm). Acest lucru îi permite să utilizeze mai multă energie din spectrele electromagnetice. R. rubrum oxidează, de asemenea, monoxidul de carbon (CO) cu hidrogen gazos ca produs final al traseului și poate utiliza sulfura la concentrații scăzute ca donator de electroni în reducerea dioxidului de carbon. În plus, este o bacterie fixatoare de azot; aceasta utilizează azotaza pentru a transforma azotul atmosferic gazos în amoniac (Munk et al, 2011).
Există mai multe aplicații ale R. rubrum în domeniul biotehnologiei. Este un sistem model pentru conversia luminii în energie chimică și pentru căile sale de fixare a azotului. De asemenea, face obiectul unor studii de rezistență la radiații. De asemenea, poate fi utilizat în mai multe moduri pentru consum. Proteobacteria poate fi o sursă de hrană pentru animale și de îngrășământ agricol. Un alt rol important în cercetare include producția de vitamine. De asemenea, este cercetată și pentru producția de plastic biologic din precursori de polihidroxibutric-acid. R. rubrum poate fi, de asemenea, un contributor în combustibilii biologici pe bază de hidrogen, în principal prin evoluția enzimei nitrogenază.
Structura genomului
Secvențierea genomului este finalizată. Acesta constă într-un cromozom circular cu 4.352.825 de perechi de baze, din care 65% sunt perechi guanină-cistină. Există, de asemenea, un plasmid cu 53.732 de perechi de baze cu 60% guanină-citosină. În total, există 3.850 de gene care codifică proteine și 83 de gene ARN. Dintre aceste gene, 6,9% sunt legate de transcripție; 4,6% de traducere, structura ribozomilor și biosinteză; 4,0% de replicare, recombinare și reparare; 7,9% de mecanisme de transducție a semnalelor; 5,9% de biogeneză a peretelui celular și a membranei; 6,6% de producere și conversie a energiei; 5,0% de transport și metabolism al carbohidraților; 9,0% de transport și metabolism al carbohidraților.9% transportul și metabolismul aminoacizilor; 4,7% transportul și metabolismul coenzimelor; 3,7% transportul și metabolismul lipidelor; și 6,5% transportul și metabolismul ionilor anorganici(Munk et al, 2011).
Ecologie
Datorită faptului că Rhodospirillum rubrum poate crește atât aerob, cât și anaerob, este capabil să locuiască într-o mare varietate de condiții. R. rubrum se găsește în multe medii acvatice naturale, cum ar fi iazuri, lacuri, cursuri de apă și ape stătătoare (Reslewic et. al, 2005). De asemenea, R. rubrum se găsește adesea în noroi și în apele reziduale (Brock et al, 2000). Studiile au arătat că R. rubrum poate face schimbări mari în compoziția sa chimică pentru a se adapta la diferite medii (Cohen-Bazire și Kunisawa, 1963). R. rubrum preferă să se dezvolte în habitate cu un pH de 6,8-7,2 (Bergey și Holt, 1994). Studiile au arătat, de asemenea, că R. rubrum are o temperatură optimă de creștere de 22-35 grade Celsius (Weaver, 1971).
Structura și metabolismul celular
R. rubrum este un organism versatil care poate obține energie printr-o varietate de mecanisme. Mecanismele de respirație și de fotosinteză există împreună, iar activitatea lor depinde de prezența luminii și a energiei. R. rubrum poate crește în medii chimio-tropicale întunecate cu prezența oxigenului sau poate crește într-un mediu fototropical fără oxigen.
Photosinteza la R. rubrum începe cu donarea unui hidrogen de la un substrat organic la un substrat oxidant. Bacterioclorofila și cartenoizii, pigmenții fotoreactivi care se găsesc în membrana celulară, sunt legați la cromatofori (Schachman, Pardee și Stanier, 1952). Cromatoforii sunt discuri aplatizate care conțin fosfolipide de colină, cardiolipină și digliceride galactosilice (Benson, Wintermans și Wiser, 1959; Benson 1961). În plus, cromatoforii conțin un lanț complet de transport al electronilor care include diverși citocromi, flavină și nucleotide de piridină (Newton și Newton, 1957; Hulcher și Conti, 1960; Kamen, 1961). Membrana celulară conține, de asemenea, mecanisme pentru sinteza ATP, inclusiv o ATP-sintetază.
În prezența oxigenului, R. rubrum este capabil să respire aerobic folosind un lanț tradițional de transport al electronilor cu NAD+/NADH ca purtător primar de electroni (Keister și Minton, 1969). Printre proteinele de transport al electronilor se numără citocromul C428 (Chance și Smith, 1955). Cu toate acestea, respirația aerobă este inhibată de fotosinteză (Oelze și Weaver, 1971).
R. rubrum este, de asemenea, capabil de respirație anaerobă. Acceptorii săi de electroni terminali neoxigenici pot include dimetilsulfoxidul și oxidul de trimetilamină. Prezența acestor acceptori de electroni face posibil ca substraturi precum succinatul, malatul și acetatul să susțină creșterea lui R. rubrum. Cu toate acestea, acești acceptori sunt doar aproximativ 33-41% la fel de eficienți ca oxigenul în ceea ce privește conservarea energiei (Schultz și Weaver, 1982).
În plus, R. rubrum are capacitatea unică de a oxida monoxidul de carbon cu ajutorul monoxidului de carbon dehidrogenază. Această cale de oxidare se termină cu reducerea și hidrogenul și producerea de hidrogen gazos.
Fără un acceptor terminal de electroni, R. rubrum finalizează fermentația acidă mixtă. Principalii produși ai fermentării piruvatului sunt acetatul, formatul, dioxidul de carbon și hidrogenul. În prezența ionului bicarbonat, fructoza este capabilă să fie fermentată. Produșii finali ai acestei căi sunt aceiași ca și în cazul piruvatului, dar cu adăugarea de succinat și propionat (Schultz și Weaver 1982).
Pe lângă capacitatea de a fixa dioxidul de carbon (Schon și Biedermann, 1972), o proprietate anabolică identificatoare a R. rubrum este capacitatea sa de a fixa azotul. În condiții de întuneric cu prezența fructozei, R. rubrum folosește azotaza pentru a fixa azotul gazos în amoniu. Conține atât azotaze cu Fe-Mo, cât și azotaze numai cu Fe.
Patologie
Nu s-a constatat că Rhodospirillum rubrum infectează oamenii sau animalele.
Current Research
Rezistența la radiații
Rhodospirillum rubrum a oferit recent informații despre rezistența la radiații. Felice Mastroleo et al, în Insight into the radiotolerance of the life support bacterium Rhodospirillum rubrum S1H by means of phenotypic and transcriptomic methods (2010), au caracterizat toleranța la radiații a R. rubrum.
Prin supunerea bacteriei la doze subletale de raze gama Cobalt-60, un tip de radiație ionizantă, au fost caracterizate în organism genele marker ale R. rubrum specifice radiațiilor ionizante. Expresia acestor gene a crescut pe măsură ce a trecut „timpul de recuperare” după un anumit tratament cu radiații gamma. Analiza expresiei genelor de către R. rubrum după radiațiile gamma creează potențialul pentru continuarea cercetărilor microbiologice care ar putea duce la progrese în ceea ce privește rezistența aplicată la radiații în viitor.
Plastice biodegradabile
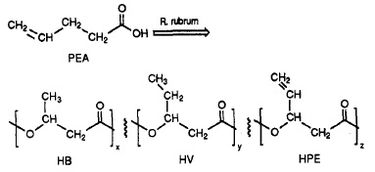
Se constată că R. rubrum produce o clasă de materiale plastice biodegradabile, poli beta-hidroxialcanoați (PHA). Aplicațiile acestei cercetări pot rezolva multe probleme cauzate de materialele plastice sintetice, nedegradabile, care au avut un impact negativ asupra mediului. Un experiment de cercetare realizat de Herbert. W. Ulmer și colab. a consemnat producerea de PHA-uri care conțin unități 3HV de către Rhodospriillum rubrum (1993). Ulmer a folosit acidul 4-pentenoic (PEA) ca mediu de creștere pentru R. rubrum. Atunci când bacteria a fost plasată în „condiții de producere a PHA”, au fost sintetizate PHA-uri care conțineau 3 subunități repetitive (notă: R = grup substituent): HB (R = metil), HV (R = etil), HPE (R = vinil) (a se vedea figura 1).
Sinteza de materiale plastice funcționale și biodegradabile de către bacterii are potențialul de a reduce poluarea globală și cheltuielile de combustibil de la fabricile de plastic. R. rubrum poate fi folosit ca instrument microbiologic pentru a produce biopolimeri ecologici.
Energie regenerabilă
Preocuparea crescândă pentru poluarea mediului cauzată de arderea combustibililor fosili a dus la o creștere a cercetărilor asupra Rhodospirillum rubrum. Utilizarea R. rubrum ca sursă de hidrogen pentru a fi utilizat în combustibilii pe bază de hidrogen regenerabil rezolvă mai multe probleme create de consumul de combustibili fosili. Hidrogenul se oxidează în apă ca produs de combustie, spre deosebire de combustibilii fosili care creează poluanți precum CO, CO2, SOx și NOx. R. rubrum poate produce hidrogen prin utilizarea următoarei „reacții de transfer de apă-gaz” (Najafpour et al., 2004):
CO + H2O → H2 + CO2
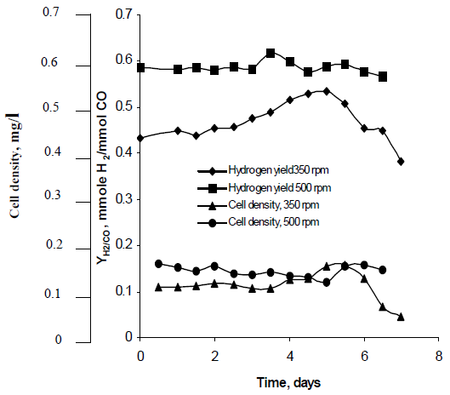
Metoda de cercetare a lui Najafpour a observat modul în care agitația, creată de seturi de rotoare de turbină setate la rate de 350 rpm, precum și de 500 rpm, a afectat ratele de producție de hidrogen ale R. rubrum. Cercetările sale au constatat că combustibilul pe bază de hidrogen are potențialul de a reduce încălzirea globală și de a oferi o soluție tehnologică ecologică pentru producția și consumul de combustibil. Studiul său a arătat că „o rată de agitare mai mare a produs o cantitate mai mare de hidrogen, precum și o densitate celulară mai mare” a R. rubrum (a se vedea figura 2).
Într-un alt studiu microbiologic, realizat de Matthew Ross Melnicki, s-a constatat că R. rubrum oferă potențialul pentru energie regenerabilă, cum ar fi combustibilul hidrogen, prin sinteza hidrogenului. Căile sale biologice pot produce niveluri de hidrogen la rate ridicate atunci când este cultivat în comun cu Chlamydomonas reinhardtii, o specie de alge cu care R. rubrum poate crește armonios. Melnicki a propus o abordare neconvențională de co-cultivare pentru sinteza hidrogenului, care a produs niveluri sporite de sinteză a hidrogenului care nu pot fi atinse în cazul în care co-cultivarea este decuplată. C. reinhardtii a fost favorizată la intensități luminoase mai mari, în timp ce R. ruburm a fost mai productivă la intensități luminoase mai mici, evidențiind avantajele co-cultivării lor.
Prin utilizarea unei tehnici de cultivare a R. rubrum pe loturi hrănite, s-a constatat că culturile inoculate au produs hidrogen în timpul fazei sale de creștere exponențială, iar sinteza hidrogenului a continuat să înflorească continuu timp de 70 de ore. Când creșterea a încetat, Melnicki a adăugat succinat, care acționează ca un donator de electroni în calea metabolică, iar evoluția hidrogenului a fost reluată, chiar dacă bacteriile nu mai creșteau acum. Cu toate acestea, este important de remarcat faptul că, odată cu adăugarea continuă de succinat, s-au constatat rate reduse de evoluție a hidrogenului. Odată ce a fost adăugat un mediu de creștere suplimentar, ratele de producere a hidrogenului au crescut din nou.
Melnicki a mai afirmat: „deși creșterea nu este necesară pentru producerea de hidrogen, această lucrare stabilește necesitatea creșterii celulelor pentru a menține ratele maxime, sugerând adecvarea industrială a unei strategii de cultură semi-continuă” (Melnicki, 2009).
Cool Factor
Secvența genomului Rhodospirillum rubrum este doar a doua din genul Rhodospirillum care a fost publicată, după R. centenum (Munk et al, 2011).

Un aspect interesant al Rhodospirillum rubrum este capacitatea sa de a-și schimba culoarea în diferite condiții. Este un fototrof anoxigen, dar nu produce oxigen extracelular. În schimb, produce sulf elementar în timpul recoltării luminii. Acest lucru se datorează faptului că R. rubrum nu are complexul 2 de recoltare a luminii (Munk et al., 2011). Bacteria utilizează bacterioclorofila, ceea ce îi permite să absoarbă lungimi de undă ale luminii pe care plantele nu le utilizează. Bacterioclorofila absoarbe lungimi de undă de 800-925 nm.
În condiții aerobe, activitatea fotosintetică a R. rubrum este suprimată genetic, iar bacteria devine incoloră. Cu toate acestea, în condiții anaerobe, fotosinteza este activată, iar bacteria capătă o culoare distinctă roșu-violet (a se vedea imaginea 5). Acest lucru se datorează activității din cromatofori, care se găsesc în membrană (Munk et al., 2011). Cromatoforii conțin pigmenții bacterioclorofilieni ai bacteriei și suferă modificări de absorbție în timpul activării fotosintezei.
Benson, A. „Chloroplast lipid metabolism”. În Simpozionul nr. VI. Al 5-lea Congres internațional de biochimie. 1961. Oxford: Pergamon Press. (In the Press.)
Benson, A., Wintermans, J. și Wiser, R. „Chloroplast lipids as carbohydrate reservoirs”. Fiziologie vegetală. 1959. Volumul 34. p. 815.
Bergey, D. și Holt, J. Bergey’s manual of determinative bacteriology. Ediția a 9-a. Baltimore. Lippincott Williams and Wilkins. 1994. p. 359.
Brock, T.D., M.T. Madigan, J.M. Martinko, și J Parker. Biology of Microorganisms. Ediția a 9-a. Upper Saddle River: Prentice Hall, 2000. p. 459.
Chance, B. și Smith, L. „Pigmenții respiratori ai Rhodospirillum rubrum”. Natura. Volume 174. p. 803-806.
Cohen-Bazire, G. și Kunisawa, R. „The Fine Structure of Rhodospirillum Rubrum .” Journal of Cell Biology. 1963. Volumul 16. p. 401-419.
Hulcher, M. și Conti, S. „Cytochronies in chlorophyll-containing particles of Chromatium and Chlorobium thiosuphatophilum”. Biochemistry and Biophysics Research Communications. 1960. Volumul 3. p. 497.
Kamen, M. „The Haem Proteins of Photosynthetic Tissues”. În Simpozionul nr. VI. Al 5-lea Congres internațional de biochimie. 1961. Oxford: Pergamon Press. (În presă.)
Keister, D. și Minton, N. „Energy-linked reactions in photosynthetic bacteria. 3. Studii suplimentare asupra reducerii nicotinamidelor-adeninelor dinucleotide legate de energie de către cromatoforii Rhodospirillum rubrum”. Biochimie. 1969. Volumul 8. p. 167-173.
Mastroleo, F., Monsieurs, P., și Leys, N. „Insight into the radiotolerance of the life support bacterium Rhodospirillum rubrum S1H by means of phenotypic and transcriptomic methods”. Adunarea științifică COSPAR. 2010. p. 4.
Melnicki, M. R. „Modes of hydrogen production in the photosynthetic bacterium, Rhodosphirillum rubrum”. University of California, Berkeley. 2009.
Munk, C., Copeland, A., Lucas, C., Lapidus, A., et al. „Complete genome sequence of Rhodospirillum rubrum type strain (S1T)”. Standarde în științele genomice. 2011. Volumul 4. pp. 293-302.
Najafpour, G., Ismail, K., Younesi, H., Mohamed, A. R., și Kamaruddin, A. H. „Hydrogen as clean fuel via continuous fermentation by anaerobic photosynthetic bacteria, Rhodospirillum rubrum”. Jurnalul african de biotehnologie. 2004. Volumul 3. pp. 503-507.
Newton, J. și Newton, G. „Composition of the photoactive subcellular particles from Chromatium”. The Archives of Biochemistry and Biophysics (Arhivele de biochimie și biofizică). 1957. Volumul 71. p. 250.
Oelze, J. și Weaver, P. „The adjustment of photosynthetically grown cells of Rhodospirillum rubrum to aerobic light conditions”. The Archives of Microbiology. 1982. Volume 79. p. 108-121.
Reslewic, S., Shigou, Z., Place, M., Zhang, Y., Briska A., Goldstein, S., Churas, C., Runnheim, R., Forrest, D., Lim, A., Lapidus, A., Han, C., Roberts, G., și Schwartz, D. „Whole-Genome Shotgun Optical Mapping of Rhodospirillum rubrum .” Microbiologie aplicată și de mediu. 2005. Volumul 71. p. 5511-5522.
Schachman, H., Pardee, A., și Stanier, R. „Studies on the macromolecular organization of microbial cells”. The Archives of Biochemistry and Biophysics (Arhivele de biochimie și biofizică). 1952. Volumul 38. p. 245-260.
Schön, G. și Biedermann, M. „Synthesis of volatile acids by fermentation of pyruvate and fructose in anaerobic dark cultures of Rhodospirillum rubrum”. Archiv fur Mikrobiologie.1972. Volumul 85. p. 77-90.
Schultz, J. și Weaver, P. „Fermentarea și respirația anaerobă de către Rhodospirillum rubrum și Rhodopseudomonas capsulate”. The Journal of Bacteriology (Jurnalul de bacteriologie). 1982. Volume 149. p. 181-190.
Ulmer, H., Gross, R., Posada, M., Weisbach, P., Fuller, R. C., și Lenz, R. „Bacterial production of poly(.beta.-hydroxyalkanoates) containing unsaturated repeating units by Rhodospirillum rubrum”. Macromolecules. 1994. Volumul 27. p. 1675-1679.
Weaver, P. „Temperature-Sensitive Mutations of the Photosynthetic Apparatus of Rhodospirillum rubrum”. Proceedings of the National Academy of the Sciences of the United States of America. 1971. Volumul 68. p. 136-138.
Editat de studenții Dr. Iris Keren: Allison Almeida, Matthew Cozzolino, Trung Nguyen și Jessica Wong.
NEUF2011
.