- Origins of Triceratops and other Ceratopsid Dinosaurs
- Variatie in kieuwen en hoorns van dezelfde soort
- The Triceratops Saga… De Grote Soorten Roundup: Van 16 soorten naar 2.
- Triceratops vs. Torosaurus – Triceratops is geen Torosaurus
- Hoorns en franjes – ze zijn niet gemaakt om te rammen!
- Triceratops Behavior: Herding and Diet
- Herding Behavior
- Tanden en Dieet
- Hoe snel kon de Triceratops Dinosaurus rennen? Triceratops en Ceratopsid Locomotion
- Triceratops Dinosaurus Huid – Schubben, Scutes, en Quills??
- Aanbevolen Tyrannosaurusboeken en Fossielen:
Origins of Triceratops and other Ceratopsid Dinosaurs

Protoceratops andrewsi specimen in het CMNH. Deze kleine, 6 voet grote ceratopsid is een van de weinige Aziatische Ceratopsid Dinosaurussen.
De Ceratopsids – Oorsprong en Verspreiding
De Ceratopsidae Dinosaurus familie omvat de viervoetige herbivoren met onderscheidende gezichtshoorns en grote franje. Tot deze dinosaurusfamilie behoort de bekende Triceratops, de eerste Ceratopsid die in 1887 werd ontdekt. Er zijn talrijke onderfamilies en geslachten van Ceratopsid. Zij worden ingedeeld naar de vorm en grootte van hun hoorns, snavels, en franje.
Ceratopsiden zijn beperkt tot Noord-Amerika en Azië (Triceratops komt alleen voor in West-Noord-Amerika). Primitieve Ceratopsiden zijn in Azië gevonden, wat erop wijst dat zij daar voor het eerst verschenen en vervolgens via een landbrug naar Noord-Amerika zijn overgestoken. Eenmaal in Noord-Amerika, diversifieerden zij snel tot hun uitsterven aan het einde van het Krijt. Er zijn ongeveer 37 geslachten van Ceratopsiden geclassificeerd. Hieronder staat een afbeelding van enkele genera van Ceratopsid-schedels, die de duidelijk verschillende vormen en afmetingen van hoorn en kieuwen laat zien.

Een afbeelding van schedels van enkele leden van de Ceratopsid Dinosaurus-familie. Let op de wilde variatie in de franje, de snavel en de hoorns. Deze afbeelding is afkomstig uit het Natural History Museum of Utah.
By skinnylawyer uit Los Angeles, California, USA , via Wikimedia Commons
Variatie in kieuwen en hoorns van dezelfde soort
Toen in het begin van de 20e eeuw Ceratopsiden werden ontdekt, werden de fragmentarische overblijfselen geclassificeerd op basis van de vorm van de kieuwen, snavels en hoorns. Er werd geen rekening gehouden met het jonge versus volwassen uiterlijk en het sexuele dimorfisme. Als gevolg daarvan werden talloze soorten geschapen.
Later, toen meer en meer fossiel bewijsmateriaal werd ontdekt, realiseerden paleontologen zich dat de schedel van vorm verandert naarmate het dier zich in verschillende ontwikkelingsstadia bevindt. Hoorns en franje waren pas op volwassen leeftijd volledig ontwikkeld (Farlow 327).
Een mooi voorbeeld hiervan is van de baby chasmosaurine ceratopsid dinosaurus die in 2013 werd ontdekt. Currie et al, publiceerden in 2016 een paper over deze babydinosaurus en vergeleken deze met volwassen chasmosarus-exemplaren (Currie et al, 2016). De franje van de baby chasmosarus ziet er heel anders uit dan die van de volwassen chasmosarus.
The Triceratops Saga… De Grote Soorten Roundup: Van 16 soorten naar 2.

Een Triceratops dinosaurusskelet tentoongesteld in het Field Museum of Natural History in Chicago
Een goed voorbeeld van het terugbrengen van het aantal Ceratopsiden zijn de Triceratops geslachten. Triceratops is de meest voorkomende dinosaurus in de Hell Creek Formatie van het westen van de Verenigde Staten. Aan het geslacht Triceratops werden ooit wel 16 soorten toegewezen op basis van alle verschillende schedelvariaties die werden gevonden. Toen paleontologen zich realiseerden dat de franje tijdens de ontwikkeling veranderde, werd Triceratops opnieuw geanalyseerd. In 1986 brachten Ostrom en Wellnhofer Triceratops terug tot slechts 1 soort, T. horridus. Zij dachten dat alle verschillen in schedel het gevolg waren van verschillende ontwikkelingsstadia en sexueel dimorfisme, en dat er niet talrijke gelijksoortige grote dieren konden zijn die allemaal in hetzelfde gebied leefden.
Iets later, in 1996, onderzocht Catherine Forester de Triceratops soorten en concludeerde, omdat T. horridus veel talrijker was dan T. prorsus, dat het twee verschillende soorten waren. Uiteindelijk, in 2009, ondersteunden Scannella en Fowler deze zienswijze en was de saga voorbij. Er waren twee Triceratops soorten… soort van… En dan is er nog die hele Torosaurus-kwestie…
Triceratops vs. Torosaurus – Triceratops is geen Torosaurus

Vergelijking van de grootte van Torosaurus latus specimens.
“A, YPM 1831, een subadult, schedel lengte >2,6 m, en B, ANSP 15192, een vroege volwassene, schedel lengte 1,8 m. Het rostrum van YPM 1831 is gereconstrueerd en zou waarschijnlijk iets langer zijn geweest, net als bij ANSP 15192.” Van: Nicholas R. Longrich*, Daniel J. Field, 2012 – PLOS one onder CC2 licentie.
Torosaurus is een grote ceratopside die in dezelfde formatie is gevonden als Triceratops. Hij is groter en de kraag is duidelijk anders. Hij is langer en heeft grote gaten aan de bovenkant. In 2010 publiceerden Scannella en Horner een artikel waarin zij aantoonden dat Torosaurus eigenlijk een laat, snel, groeistadium van Triceratops was. Dit deed allerlei controverses oplaaien. Paleontologen kozen partij, sommigen waren het eens en sommigen oneens. Persoonlijk heb ik er altijd moeite mee gehad hoe twee grote gaten in het kieuwbeen konden eroderen. Dat zou werkelijk de hele spieraanhechtingsstructuur veranderen. De laatste jaren zijn er veel artikelen verschenen die de resultaten van Scannella en Horner weerleggen. In 2011 publiceerde de ceratopside dinosaurusexpert Dr. Andy Farke een artikel waarin hij aantoonde dat Torosaurus “een geslacht is dat zich onderscheidt van Triceratops en Nedoceratops”. Recente artikelen lijken het aparte geslacht te ondersteunen, waaronder een artikel van Longrich en Field in 2012. Zij bestudeerden verschillende Torosaurus en Triceratops schedels en deden een clustering analyse om ze in groeireeksen te rangschikken. Zij vonden onvolwassen en volwassen individuen van zowel Torosaurus als Triceratops. Daarom zouden zij hetzelfde dier kunnen zijn. Triceratops en Torosaurus zijn verschillende dinosaurussen.
De afbeelding hierboven is van Longrich en Field (2012). Het toont een volwassen en een subadulte Torosoarus.
Hoorns en franjes – ze zijn niet gemaakt om te rammen!

Schedel van een dinosaurus uit Utah met uitgebreide hoorns en kieuwen, tentoongesteld in het CMNH
De meeste Ceratopsiden hadden enorme kieuwen, met uitzondering van de Diceratops en de Triceratops. Deze twee genera hadden kieuwen die naar verhouding kleiner waren (Fastovsky blz. 180). Toename van de grootte van de kieuwen tijdens het leven had direct invloed op de sterkte van de kaakspieren (hoe groter de kieuwen, hoe groter/sterker de kaakspieren) (Fastovsky p. 186). De grote gaten die in sommige kieuwen te zien zijn, werden waarschijnlijk opgevuld met spieraanhechtingen en/of kraakbeen.
Wat de hoorns betreft, vergeten veel mensen bij het zien van een fossiele Triceratops of andere Ceratopside Dinosauriërs, dat de hoorns op de schedel de botkernen zijn. Deze botkernen zouden zijn bedekt door de eigenlijke hoorns, die bij leven veel langer zouden zijn geweest.
Het is helaas onmogelijk om na te gaan waarvoor hoorns en franje werkelijk werden gebruikt. Maar als we moderne kuddedieren als analogie gebruiken, is het waarschijnlijk dat ze werden gebruikt voor dominantie, verdediging en vertoon. Ceratopsiden kunnen hun hoorns binnen hun eigen soort hebben gebruikt voor territoriumdrift en paringsrechten. Dit komt overeen met het fossiele bewijsmateriaal, want in schedels en franje van Triceratops zijn vaak steekwonden gevonden van andere Triceratops, wat wijst op een vorm van intraspecies gevecht (Farke, et al, 2009).
Een studie toonde aan dat als twee volwassen Triceratops elkaar frontaal zouden rammen, hun schedels zouden versplinteren! De hoorns van Ceratopsiden hadden echter een andere vorm dan die van zoogdieren. In plaats van te rammen, hebben ze misschien gewoon de hoorns op elkaar geklemd en gevochten. Een studie met behulp van modellen toonde aan dat het fysiek mogelijk was voor triceratops om hoorns op slot te houden (Farke, Andrew, A. 2004). Zij vonden 3 mogelijke hoorn-sluitingen. In plaats van elkaar te rammen, zouden zij de hoorns vastgrijpen en vechten, ongeveer zoals elanden de hoorns vastgrijpen wanneer zij vechten.
Het is interessant op te merken, dat er één uitzondering is. Een ander geslacht, Centrosaurus, een ceratopside met slechts 1 grote neushoorn, werd bestudeerd door Farke (Farke, et al, 2009) en bleek geen verwondingen te hebben. Dit betekent dat Centrosaurussen hun hoorns en franje waarschijnlijk niet gebruikten om te paren of voor territoriale gevechten, maar in plaats daarvan om te pronken, zoals een pauw. Een figuur uit Farke’s artikel is hieronder afgebeeld. Het toont de incidentie van letsels in (A) Triceratops en (B) Centrosaurus.
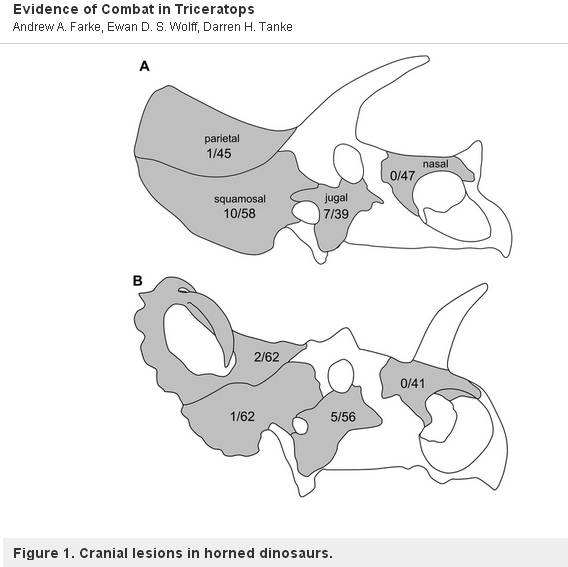
“Schema’s van de schedels van (A) Triceratops en (B) Centrosaurus, die de incidentie van letsels tonen (periosteaal reactief bot en fractuur eelt) op elk schedelelement (aantal abnormale elementen / totaal aantal elementen). Niet op schaal.”
Van: Andrew A. Farke, Ewan D. S. Wolff, Darren H. Tanke, 2009
– PLOS one under CC2 license.
Moraal van het verhaal, als Triceratops ze gebruikte voor paringsgedrag en Centrosaurus ze gebruikte voor vertoning, werden hoorns en franje waarschijnlijk op allerlei verschillende manieren gebruikt door verschillende soorten ceratopsiden.
Eén gebruik van de hoorns en franje door veel ceratopsiden was waarschijnlijk om roofdieren af te weren. Sommige ceratopside fossielen zijn zelfs gevonden met genezen T-rex bijtwonden, dit zijn exemplaren die met succes een T-Rex aanval hebben afgeweerd. Terwijl vele andere gevonden zijn met niet-genezen bijtwonden, wat betekent dat ze T-Rex voedsel waren. Vanwege deze fossiele vondsten is het niet ondenkbaar om aan te nemen dat de franje en hoorns ook werden gebruikt om de machtige T-Rex af te weren die in het late Krijt in het westen van de V.S. leefde.
Of hoorns en franje nu werden gebruikt voor hoornafsluiting, hoofdgors, vertoon, verdediging, of al het bovenstaande, één ding is zeker. Ze waren er in alle vormen en maten, kijk maar naar onderstaande afbeelding!

Ceratopsid Dinosaur hoorns en franje te zien in het Carnegie Museum of Natural History.Let op de eerste schedel links (Pachyrhinosaurus sp.) heeft helemaal geen hoorn, in plaats daarvan een dikke botknobbel.
Triceratops Behavior: Herding and Diet
Herding Behavior
In het Midwesten zijn grote botbedden gevonden met honderden Centrosaurusfossielen, die erop wijzen dat ze in grote kuddes reisden. Van Triceratops zijn geen grote botbeddingen gevonden, maar er zijn wel groepen individuen bij elkaar gevonden, zodat het mogelijk is dat zij in grote kudden of in ieder geval in kleinere groepen reisden. Dit is in overeenstemming met het feit dat hoorngebruik vaker voorkomt bij kuddedieren, zoals herten, gazellen en buffels.

Afbeelding van drie Triceratops-tanden, gevonden in de Hell Creek Formatie van South Dakota tijdens mijn Hell Creek Dinosaur Dig.
Tanden en Dieet
Zoals alle kuddedieren waren Ceratopsiden herbivoren. Hun bek was bij uitstek geschikt voor het eten van taaie vegetatie. Deze dinosauriërs hadden tanden met dubbele wortels die waren gerangschikt in groepen die tandbatterijen werden genoemd. Een tandbatterij is een unieke tandopstelling waarbij aangrenzende tanden in rijen in de lengterichting en in verticale kolommen aan elkaar vastzaten. Met ten minste 3 tanden in elke kolom, 1 was functioneel en 2 waren “vervangtanden”. De tanden, die tijdens het leven van de dinosauriërs voortdurend werden vervangen, waren zo verticaal geplaatst en waren zo scherp (zelfs slijpend) dat ze als een schaar konden knippen. Tijdens het kauwen raakten de tanden van deze dinosaurussen elkaar helemaal niet zoals bij de mens; in plaats daarvan gleden ze langs elkaar heen, in een snijdende beweging (Fastovsky 183). Vanwege deze vlijmscherpe tanden en zeer sterke kaakspieren, worden zij vaak “Nature’s first Cuisinarts!!!” genoemd. (Farlow 85). Zij konden door het hardste materiaal snijden.
Terzijde, de andere veel voorkomende planteneters, de Hadrosaurs, hadden soortgelijke tandbatterijen.
Wetenschappers zijn enigszins onzeker over hun exacte plantendieet. Zij vermoeden dat zij, door hun relatief geringe snuffelhoogte, laag groeiende planten zoals angiospermen, varens en kleine coniferen die op die hoogte groeiden, gegeten kunnen hebben. Door hun grote en zware schedels konden ze niet op hun achterpoten staan, maar deze grasmaaiers van het Amerikaanse westen waren misschien wel in staat om over bomen heen te bulldozeren om zo bij de bladeren te komen.

Afbeelding van een Triceratops-tand die tijdens mijn Hell Creek Dinosaur Dig.
Hoe snel kon de Triceratops Dinosaurus rennen? Triceratops en Ceratopsid Locomotion
Velen gaan ervan uit dat de Triceratops een zeer traag en log dier was. Het bewijs toont aan dat dit niet het geval is.
Skeletreconstructies van Ceratopsid Dinosaurussen in vele musea (inclusief de in dit artikel afgebeelde exemplaren) tonen Triceratops met een uitgestrekte hagedisachtige houding. Onderzoek toont aan dat deze dinosaurussen een meer zoogdierachtige houding zouden moeten hebben, met de voorpoten onder het dier, niet zijwaarts uitgespreid. De hagedisachtige houding is het gevolg van fouten bij het monteren van de ribben en wervels van Ceratopsiden.
Het bewijs voor bovenstaande beweringen komt van analyses van Triceratops en andere Ceratopsische sporen. Paul en Christiansen publiceerden in 2000 een artikel waarin zij de pedaal indrukken in sporen bestudeerden en de plaatsing en de hoeken van de handen en voeten analyseerden. Op basis van deze informatie stelden zij vast dat Ceratopsiden een olifantachtige voorpoot houding hebben. Een zoogdierachtige houding stelt Ceratopsiden in staat om VEEL sneller te lopen dan een reptielachtige houding. Paul en Christiansen geven vervolgens aan dat de maximale loopsnelheid van de grootste Ceratopsiden “in grote lijnen overeenkomt met die van neushoorns”. Tussen haakjes, de zwarte neushoorn kan rennen met snelheden van rond de 55 km/u, of 34 mijl per uur! Dat is behoorlijk snel!

Een Triceratops skeletreconstructie van het Carnegie Museum of Natural History toont een uitgestrekte, hagedisachtige houding, wat volgens recent bewijsmateriaal niet correct is.

Een Triceratops model van het Field Museum in Chicago toont een meer zoogdierachtige houding, wat volgens recent bewijsmateriaal wel correct is.
Een Triceratops model van het Field Museum in Chicago toont een meer zoogdierachtige houding, wat volgens recent bewijsmateriaal wel correct is.
Triceratops Dinosaurus Huid – Schubben, Scutes, en Quills??
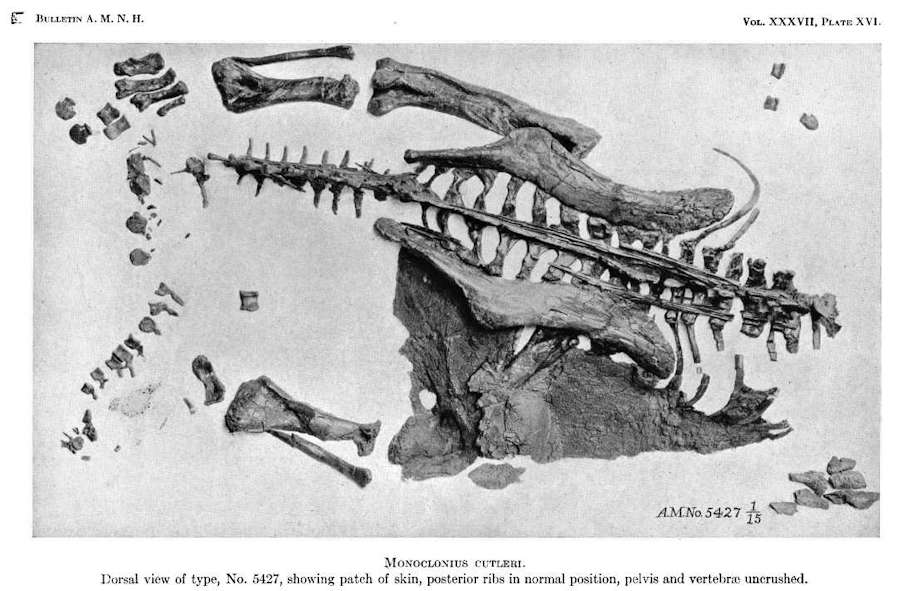
Afbeelding van een Centrosaurus dinosaurus specimen (AMNH 5427) compleet met een grote huidlap
Datum: 1917, Auteur: Barnum Brown,Bron: AMNDH Digitale Bibliotheek: B037a10.pdf Public Domain.
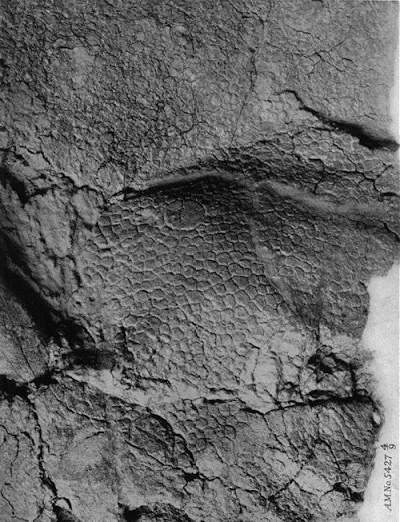
Closeup van de AMNH 5427 Centrosaurus dinsoaurus huidafdruk.
Datum: 1917, Auteur: Barnum Brown,Bron: B037a10.pdf Public Domain.
Gefossiliseerde Dinosaurus huidimpressies zijn ZEER zeldzaam om te vinden. Men noemt Dinosaurusfossielen met huid vaak “Dino Mummies”, hoewel ze niet gemummificeerd zijn. De eigenlijke huid is niet bewaard gebleven, maar indrukken van de huid in de ooit zachte modder die het dier omringde fossiliseren.
Gelukkig voor liefhebbers van Triceratops, zijn er een paar “Ceratopsid Mummies” gevonden. Een van de originele, hierboven afgebeeld, AMNH 5427, is een Centrosaurus waarvan een grote lap huid bewaard is gebleven.
Een recentere “Triceratops Mummie” met de naam Lane werd in 2002 in Wyoming ontdekt. Het Black Hills Institute of Geological Research heeft het T. horridus specimen zorgvuldig en nauwgezet opgegraven. Het is nu te zien in hun museum samen met een ander bijna volledig intact T. horridus skelet genaamd Kelsey.
De Triceratops Lane heeft de meest bewaarde huid dan enig ander gevonden dinosaurusfossiel. Er zijn gebieden waar meerdere voeten huid bewaard zijn gebleven.
De huid is heel anders dan veel paleontologen dachten. De onderkant van het dier is bedekt met scutes, vergelijkbaar met een krokodil. De rest van de dinosaurus is bedekt met kleine schubben. Er zijn echter ook onregelmatige schubachtige vormen. Sommige zijn vuistvormige zeshoeken en hebben kleine porieachtige gaatjes in het midden.
Sommigen hebben gespeculeerd dat uit deze gaten stekels kunnen zijn gestoken. Deze “Quill” hypothese is afkomstig van een verwant van de Triceratops, de Psittacosaurus uit Azië. Het is een ver familielid dat stekelachtige structuren op de top van zijn staart heeft. Het is belangrijk op te merken dat Psittacosaurur een ver familielid van Triceratops is, dus dat betekent niet dat Triceratops ook stekels zou hebben gehad. De “Quill” hypothese ligt momenteel in het rijk van speculatie, en we zullen moeten wachten op verdere studies en publicaties die licht werpen op de exacte aard van Triceratops huid. Hieronder is een afbeelding te zien van een doorsnede van de huid van de Triceratops Lane.

Doorsnede van de huid van de Triceratops, afkomstig van de dinosaurus Lane, waarop de zeshoekige schubben te zien zijn. Afbeelding door:Black Hills Institute of Geological Research

Pittacosaurus-dinosaurus met bewaard gebleven dekveren en een model. Dit is een verre verwant van Triceratops. Paleontologen speculeren dat de stekelstructuren uit sommige van de oneven schubben van Triceratops zouden kunnen zijn gestoken.
Aanbevolen Tyrannosaurusboeken en Fossielen:

New Perspectives on Horned Dinosaurs: The Royal Tyrrell Museum Ceratopsian Symposium (Life of the Past)
Dit boek geeft een geactualiseerde kijk op ceratopsians. Het presenteert nieuw onderzoek naar de functies, biologie, gedrag, paleoecologie en paleogeografie van gehoornde dinosaurussen. Het beschrijft ook vele nieuwe soorten. Het boek is een verzameling van wetenschappelijke artikelen, dus dit is alleen voor serieuze dinosaurus lezers! Ik denk dat het een geweldig naslagwerk is en een must bron voor dinosaurus liefhebbers.

Dinosaurs Rediscovered: The Scientific Revolution in Paleontology
door Michael J. Benton, 2019
Dit is een geweldig algemeen boek over dinosaurussen. Het is up-to-date en gaat over de nieuwe ontdekkingen van de afgelopen 20 jaar die onze kijk op dinosaurussen volledig heeft veranderd. Het boek is opgedeeld in makkelijke hoofdstukken die ingaan op hun kleuren, snelheden, bijtkracht, kinderverzorging, en meer… dingen die we jaren geleden nog niet wisten! Bekijk de recensies en de voorbeeldpagina’s.

Hoogwaardige dinosaurustanden van Fossilera
Referenties / geciteerde werken
Currie Philip J., Holmes Robert B., J. Ryan Michael, Coy Clive. (2016) Een juveniele chasmosaurine ceratopside (Dinosauria, Ornithischia) uit de Dinosaur Park Formation, Alberta, Canada. Journal of Vertebrate Paleontology, 2016; e1048348 DOI: 10.1080/02724634.2015.1048348
Farlow, O. James, Brett-Surman, M.K., Editors (1997). De complete dinosaurus. IN: Indiana University Press.
Fastovsky, E. David, Weishampel, B. David (1996). The Evolution And Extinction Of The Dinosaurs. NY: Cambridge University Press.
Farke A.A. (2011) Anatomy and Taxonomic Status of the Chasmosaurine Ceratopsid Nedoceratops hatcheri from the Upper Cretaceous Lance Formation of Wyoming, U.S.A. PLoS ONE 6(1): e16196. doi: 10.1371/journal.pone.0016196
Farke AA, Wolff EDS, Tanke DH. (2009) Bewijs van vechten bij Triceratops. PLoS ONE 4(1): e4252. doi: 10.1371/journal.pone.0004252
Farke, A.A. 2004. Hoorngebruik bij Triceratops (Dinosauria: Ceratopsidae): Het testen van gedragshypothesen met behulp van schaalmodellen. Palaeontologia Electronica 7(1):10 pp. (PDF)
Forster Catherine A. (1996) Soortenresolutie bij Triceratops: cladistische en morfometrische benaderingen. Journal of Vertebrate Paleontology, 16:259-270.
Longrich NR, Field DJ. (2012) Torosaurus is geen Triceratops: Ontogeny in Chasmosaurine Ceratopsids as a Case Study in Dinosaur Taxonomy. PLoS ONE 7(2): e32623. doi:10.1371/journal.pone.0032623
Ostrom J.H, Wellnhofer P. (1986) The Munich specimen of Triceratops with a revision of the genus. Zitteliana. 14, 111-158.
Scannella J, Horner JR. (2010) Torosaurus is Triceratops, synonymie door ontogenie. Journal of Vertebrate Paleontology 30: 1157-1168.
Scannella, J.B. and D.W. Fowler. (2009) Anagenese bij Triceratops: bewijs uit een nieuw opgelost stratigrafisch raamwerk voor de Hell Creek Formation. North American Paleontological Convention , abstracts volume: 148-9.
Gregory S. Paul, Per Christiansen. (2000) Forelimb posture in neoceratopsian dinosaurs: implications for gait and locomotion Paleobiology Summer, v. 26 no. 3 p. 450-465