Rhodospirillum rubrum属に関する微生物バイオレームページ
分類
高次分類
界: Bacteria
Phylum: Proteobacteria
Class: Alphaproteobacteria
Order: Rhodospirillales
Family: ロードスピリル科
属 Rhodospirillum
種
Rhodospirillum rubrum
性状と意義
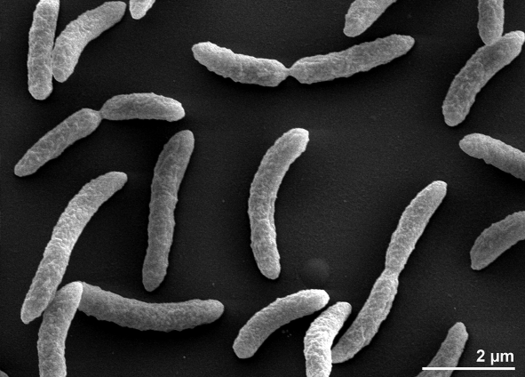
Rhodospirillum rubrumはグラム陰性の中好性プロテオバクテリアで、その名前は「Rhodospirillum」です。 最適な生育温度は25~30℃です。 多層の外被で、細胞壁の大部分は不飽和脂肪酸ですが、一部飽和脂肪酸も含んでいます。 R. rubrumはスピリラ(spirilla)、つまり螺旋状の形をしている。 極性鞭毛を持つため、運動性がある。 長さは3-10μm、幅は0.8-1.0μm。
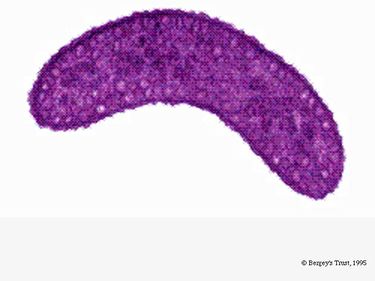
R. rubrumは通性嫌気性細菌である。 酸素の有無によって、アルコール発酵や好気性呼吸を行うことができる。 また、光合成を行うことができ、色素体粒子にカロテノイドとバテリオクロロフィルを含んでいます。 これらの分子は光を吸収してエネルギーに変換するのに役立ち、また嫌気状態では赤紫色の独特の色を呈する。 好気的条件下では無色である。
光合成は好気的条件下では活発だが、酸素が存在すると一般に抑制される。 光合成の主な副産物はO2ではなく、硫黄である。 R. rubrumは光合成を行う場合、従属栄養的に成長することも、自給的に成長することも可能です。 多くの植物と異なり、R. rubrumはクロロフィルaを含まない(吸収スペクトル430-662 nm)。 しかし、クロロフィルb(吸収波長660-680 nm)とバクテリオクロロフィル(吸収波長800-925 nm)は持っている。 そのため、電磁波スペクトルのエネルギーをより多く利用することができる。 また、R. rubrumは一酸化炭素(CO)を酸化して水素ガスを経路の最終生成物とし、低濃度の硫化物を二酸化炭素の還元における電子供与体として利用することができる。 さらに、窒素固定菌でもあり、ニトロゲナーゼを用いて大気中の窒素ガスをアンモニアに変換します(Munk et al, 2011)。
R. rubrumのバイオテクノロジー分野での応用はいくつかあります。 光から化学エネルギーへの変換や、窒素固定経路のモデル系となっています。 また、耐放射線性の研究対象にもなっている。 消費用としても、いくつかの方法で利用することができる。 プロテオバクテリアは、動物の餌や農業用肥料の原料になる可能性があります。 研究上のもう一つの重要な役割として、ビタミンの生産があります。 また、ポリヒドロキシブトリック酸の前駆体からバイオプラスチックを生産することも研究されている。 R. rubrumは、主にニトロゲナーゼという酵素を進化させることで、生物学的水素燃料の分野でも貢献できるかもしれない。 4,352,825塩基対の円形染色体からなり、その65%はグアニン-シストシン対である。 また、53,732塩基対のプラスミドがあり、その60%がグアニン-シトシン対である。 合計で、3,850個のタンパク質コード遺伝子と83個のRNA遺伝子がある。 これらの遺伝子のうち、転写関連は6.9%、翻訳・リボソーム構造・生合成は4.6%、複製・組換え・修復は4.0%、シグナル伝達メカニズムは7.9%、細胞壁・膜生合成は5.9%、エネルギー生産・変換は6.6%、糖質輸送・代謝は5.0%、9.9%、補酵素の輸送と代謝4.7%、脂質の輸送と代謝3.7%、無機イオンの輸送と代謝6.5%(Munk et al, 2011)
Ecology
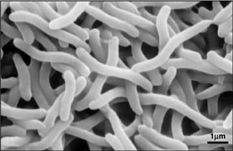 Image 3 – Rhodospirillum rubrum, SEM. (Copyright Regents of the University of California)
Image 3 – Rhodospirillum rubrum, SEM. (Copyright Regents of the University of California)Rhodospirillum rubrumは好気性にも嫌気性にも成長できるため、さまざまな環境に生息することができます。 R. rubrumは、池、湖、小川、立水など、多くの自然水系環境に生息しています(Reslewic et.) また、泥や汚水中にも多く生息しています(Brock et al, 2000)。 R. rubrumは異なる環境に適応するために化学組成を大きく変化させることができることが研究で示されている(Cohen-Bazire and Kunisawa, 1963)。 R. rubrum は pH6.8-7.2 の環境を好んで生育する(Bergey and Holt, 1994)。 また、R. rubrumの最適生育温度は22-35℃であることが研究で示されている(Weaver, 1971)。
細胞の構造と代謝
R. rubrumは様々なメカニズムでエネルギーを得ることができる多才な生物である。 呼吸機構と光合成機構は共に存在し、その活動は光とエネルギーの存在に依存する。 R. rubrumは酸素のある暗い化学熱帯環境でも、酸素のない光熱帯環境でも成長できます。
R. rubrumの光合成は、有機基質から酸化基質への水素の供与で始まります。 細胞膜に存在する光反応性色素であるバクテリオクロロフィルとカルテノイドは色素体に結合している(Schachman, Pardee and Stanier, 1952)。 色素胞は扁平な円盤状で、コリンリン脂質、カルジオリピン、ガラクトシルジグリセリドを含む(Benson, Wintermans, and Wiser, 1959; Benson 1961)。 さらに、色素体は、様々なチトクロム、フラビンおよびピリジンヌクレオチドを含む完全な電子輸送チェーンを含む(Newton and Newton、1957; Hulcher and Conti、1960; Kamen、1961)。 また、細胞膜にはATP合成酵素を含むATP合成機構が存在する。
R. rubrumは酸素存在下では、NAD+/NADHを主要電子キャリアとする従来の電子輸送系を用いて好気的呼吸ができる(Keister and Minton, 1969)。 電子輸送タンパク質の中には、シトクロムC428がある(Chance and Smith, 1955)。 しかし,好気的呼吸は光合成によって阻害される(Oelze and Weaver, 1971)。
R. rubrumは嫌気的呼吸も可能である。 その非酸素的な末端電子受容体にはジメチルスルホキシドやトリメチルアミンオキシドが含まれることがある。 これらの電子受容体の存在により、コハク酸、リンゴ酸、酢酸などの基質がR. rubrumの成長をサポートすることが可能になる。 しかし、これらの電子受容体は、エネルギー保存の点では酸素の約33-41%しか効率がない(Schultz and Weaver, 1982)。
さらに、R. rubrumは一酸化炭素脱水素酵素を使って一酸化炭素を酸化する独自の能力も持っている。 この酸化経路は、水素の還元と水素ガスの生産で終わる。
末端電子受容体がない場合、R. rubrumは混合酸発酵を完了する。 ピルビン酸発酵の主な生成物は、酢酸、ギ酸、二酸化炭素、水素である。 重炭酸イオンの存在下では、フルクトースも発酵させることができる。 この経路の最終生成物はピルビン酸のものと同じであるが、コハク酸とプロピオン酸が加わる(Schultz and Weaver 1982)。
二酸化炭素を固定する能力に加えて、R. rubrumの同化特性は窒素を固定する能力である (Schon and Biedermann, 1972)。 フルクトースの存在する暗黒条件下で、R. rubrumはニトロゲナーゼを用いて窒素ガスをアンモニウムに固定する。 4851>
Pathology
R. rubrumはヒトや動物への感染は見つかっていない。
最近の研究
放射線耐性
Rhodospirillum rubrumは最近放射線耐性について洞察を与えてくれるようになった。 Felice Mastroleoらは、Insight into the radiotolerance of the life support bacterium Rhodospirillum rubrum S1H by means of phenotypic and transcriptomic methods (2010) において、R. rubrumの放射線耐性を特徴づけています。
電離放射線であるコバルト60ガンマ線を亜致死量に曝露して、電離放射線に対する特異的なR. Rubrumマーカー遺伝子が菌内で特徴づけられていることが確認されたのです。 これらの遺伝子は、ガンマ線照射後の「回復時間」が経過するにつれて発現が増加することがわかった。 ガンマ線照射後のR. rubrumによる遺伝子発現の解析は、将来、応用放射線抵抗性の進歩につながる可能性のある、さらなる微生物学的研究の可能性を生み出します。
生分解性プラスチック
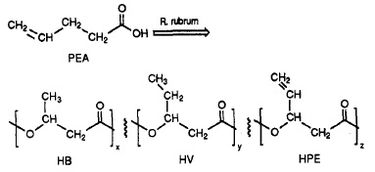
R. rubrum が生分解性プラスチック、ポリ ベータ ヒドロキシ アルカン酸塩 (PHA) クラスを生成することが判明しました。 この研究の応用は、環境に悪影響を与えている合成の非分解性プラスチックが引き起こす多くの問題を解決することができます。 ハーバートが行った研究実験。 W. Ulmer らは、Rhodospriillum rubrum による 3HV ユニットを含む PHA の生産を記録している(1993)。 Ulmerは、R. rubrumの増殖培地として4-ペンテノイック酸(PEA)を使用した。 この細菌を「PHA 産生条件」に置くと、HB(R = メチル)、HV(R = エチル)、HPE(R = ビニル)の 3 つの繰り返しサブユニット(注:R = 置換基)を持つ PHA を合成した(図 1 参照)。
バクテリアによる機能的な生分解性プラスチックの合成は、プラスチック工場による地球規模の汚染と燃料消費を減らす可能性があります。 R. rubrumは環境に優しいバイオポリマーを生産する微生物学的ツールとして利用できます。
再生可能エネルギー
化石燃料の燃焼による環境汚染への関心が高まり、Rhodospirillum rubrumの研究が盛んになりました。 R. rubrumを再生可能な水素燃料に使用するための水素源として使用することで、化石燃料の消費によって生じるいくつかの問題を解決することができます。 化石燃料はCO、CO2、SOx、NOxなどの汚染物質を発生させるが、水素は燃焼生成物として酸化して水になる。 R. rubrumは、以下のような「水-ガスシフト反応」を利用して、水素を生産することができる(Najafpour et al, 2004)。
CO + H2O → H2 + CO2
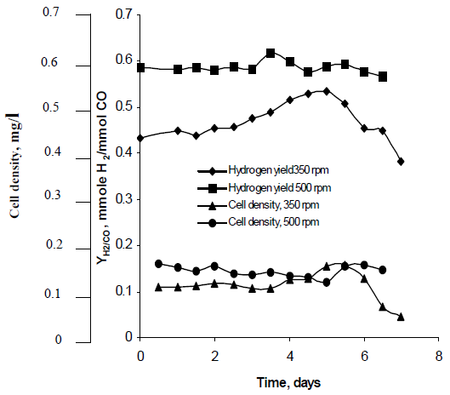
Najafpour の研究方法は、350 rpm と 500 rpm にセットしたタービン羽根車による撹拌が、ラムの水素生成率にどのように影響を与えるかを観察しています。 その結果、水素燃料は地球温暖化を抑制し、燃料生産と消費における環境にやさしい技術的解決策となる可能性があることがわかった。 彼の研究は、R. rubrum の「より高い撹拌速度が、より高い水素と細胞密度をもたらした」ことを明らかにしました (図 2 参照)。
Matthew Ross Melnicki による別の微生物学研究において、R. rubrum が水素合成によって水素燃料などの再生可能エネルギーの可能性を秘めていることが判明しました。 その生物学的経路は、R. rubrumが調和して生育できる藻類であるChlamydomonas reinhardtiiと共培養すると、高い割合で水素レベルを生成することができます。 Melnickiは、水素を合成するための従来にない共培養方法を提案し、共培養が非連結である場合には達成できないかもしれない水素合成レベルの向上を実現した。 C. reinhardtiiは高い光強度で、R. ruburmは低い光強度でより生産性が高く、両者の共培養の利点が強調されました。
R. rubrumの培養に供給バッチ法を用いることで、植え付けた培養物が指数増殖期で水素を生成し、70時間連続で水素合成が盛んであることが分かりました。 成長が止まったところで、代謝経路の電子供与体であるコハク酸を添加すると、菌は成長しなくなったが、水素発生が再開された。 しかし、コハク酸を添加し続けると、水素の発生速度が低下することが判明した。 また、Melnicki は、「水素生成に成長は必要ありませんが、この研究は、最大速度を維持するために細胞の成長の必要性を立証しており、半連続培養戦略の産業への適合性を示唆しています」と述べています (Melnicki, 2009)。
Cool Factor
Rhodospirillum rubrumのゲノム配列は、R. centenum (Munk et al, 2011) に次いでRhodospirillum属で2番目に発表された。

Rhodospirillum rubrumの興味深い点は、異なる条件で色を変えることができる点である。 無酸素性光合成細菌ですが、細胞外の酸素は生産しません。 その代わり、光を吸収する際に元素状硫黄を生成する。 これは、R. rubrumが光捕集複合体2を持たないためである(Munk et al, 2011)。 この細菌は、バクテリオクロロフィルという物質を用いて、植物が利用しない波長の光を吸収することができる。 バクテリオクロロフィルは800-925 nmの波長を吸収する。
好気的な条件下では、R. rubrumの光合成活性は遺伝的に抑制され、細菌は無色になる。 しかし、嫌気性条件下では光合成が活性化し、菌体は赤紫色を呈します(図5参照)。 これは、膜に存在する色素体(クロマトフォア)の活動によるものだ(Munk et al, 2011)。 クロマトフォアには細菌のバクテリオクロロフィル色素が含まれており、光合成活性化時に吸収変化を起こす。
Benson, A. “Chloroplast lipid metabolism.”. シンポジウムNo. VI. 5th international Congress on Biochemistry. 1961. Oxford: Pergamon Press. (In the Press.)
Benson, A., Wintermans, J. and Wiser, R. “Chloroplast lipids as carbohydrate reservoirs.”(葉緑体脂質は炭水化物の貯蔵庫). 植物生理学. 1959. 4851>
Bergey, D. and Holt, J. Bergey’s Manual of Determinative Bacteriology.「決定論的細菌学のマニュアル」. 第九版. ボルチモア Lippincott Williams and Wilkins. 1994. p. 359.
Brock, T.D., M.T. Madigan, J.M. Martinko, and J Parker.日本学術振興会特別研究員(PD)。 微生物の生物学. 第9版. アッパーサドルリバー: Prentice Hall, 2000. p. 459.
Chance, B. and Smith, L. “Rhodospirillum rubrumの呼吸色素” Nature. Volume 174. p. 803-806.
Cohen-Bazire, G. and Kunisawa, R. “The Fine Structure of Rhodospirillum Rubrum .”. ジャーナル・オブ・セル・バイオロジー. 1963. 第16巻. p. 401-419.
Hulcher, M. and Conti, S. “Cytochronies in chlorophyll-containing particles of Chromatium and Chlorobium thiosuphatophilum.”. 生化学・生物物理学研究通信. 1960. 第3巻. p. 497.
Kamen, M. “The Haem Proteins of Photosynthetic Tissues.”(光合成組織のヘムタンパク質). シンポジウムNo. VI. 第5回国際生化学会議. 1961. オックスフォード: パルガモン プレス. (In the Press.)
Keister, D. and Minton, N. “Energy-linked reactions in photosynthetic bacteria. 3. Rhodospirillum rubrum色素体によるエネルギー連鎖型ニコチンアミド・アデニンジヌクレオチド還元に関するさらなる研究”. バイオケミストリー。 1969. このような状況下において、「生命維持細菌Rhodospirillum rubrum S1Hの放射線耐性に関する表現型およびトランスクリプトーム法による洞察」(日本学術振興会特別研究員、日本学術振興会特別研究員、日本学術振興会特別研究員)、「生命維持細菌の放射線耐性に関する表現型およびトランスクリプトーム法による洞察」(日本学術振興会特別研究員)。 COSPAR Scientific Assembly. 2010. p. 4.
Melnicki, M. R. “Modes of hydrogen production in the photosynthetic bacterium, Rhodosphirillum rubrum”. カリフォルニア大学バークレー校。 2009.
Munk, C., Copeland, A., Lucas, C., Lapidus, A., et al. “Complete genome sequence of Rhodospirillum rubrum type strain (S1T)”. ゲノム科学における標準。 2011. 4巻 293-302.
Najafpour, G., Ismail, K., Younesi, H., Mohamed, A. R., and Kamaruddin, A. H. “Hydrogen as clean fuel via continuous fermentation by anaerobic photosynthetic bacteria, Rhodospirillum rubrum”. “嫌気性光合成細菌による水素の連続発酵”.Najafpour, G., Ismail, K., H., モハメド、 A. R., and カマルディン、A. H. アフリカン・ジャーナル・オブ・バイオテクノロジー。 2004. Volume 3. pp. 503-507.
Newton, J. and Newton, G. “Composition of the photoactive subcellular particles from Chromatium”(クロマティウムの光活性細胞粒子の組成). 生化学と生物物理学のアーカイブス. 1957. 4851>
Oelze, J. and Weaver, P. “The adjustment of photosynthetically grown cells of Rhodospirillum rubrum to aerobic light conditions. “光合成で成長したRhodospirillum rubrumの好気的光条件への適応。 ザ・アーカイブス・オブ・マイクロバイオロジー。 1982. このような背景のもとで、私たちは、「環境と調和した生物多様性の保全と持続可能な利用」を目指し、「環境と調和した生物多様性の保全と持続可能な利用」に取り組んでいます。 Applied and Environmental Microbiology. 2005. 第 71 巻, p. 5511-5522.
Schachman, H., Pardee, A., and Stanier, R. “Studies on the macromolecular organization of microbial cells.”(微生物細胞の高分子組織に関する研究)。 生化学と生物物理学のアーカイブス. 1952. Volume 38. p. 245-260.
Schön, G. and Biedermann, M. “Synthesis of volatile acids by fermentation of pyruvate and fructose in anaerobic dark cultures of Rhodospirillum rubrum. “嫌気性暗所培養によるピルビン酸の発酵. Archiv fur Mikrobiologie.1972. Volume 85. p. 77-90.
Schultz, J. and Weaver, P. “Fermentation and anaerobic respiration by Rhodospirillum rubrum and Rhodopseudomonas capsulate.”. ジャーナル・オブ・バクテリオロジー。 1982. 4851>
Ulmer, H., Gross, R., Posada, M., Weisbach, P., Fuller, R. C., and Lenz, R. “Bacterial production of poly(.beta.-hydroxyalkanoates) containing unsaturated repeating units by Rhodospirillum rubrum”.日本学術振興会特別研究員(PD)。 Macromolecules. 1994. 第27巻 p. 1675-1679.
Weaver, P. “Temperature-Sensitive Mutations of the Photosynthetic Apparatus of Rhodospirillum rubrum”(ロドスピリラムの光合成装置の温度感受性突然変異)。 アメリカ国立科学アカデミー紀要. 1971. Volume 68. p. 136-138.
Edited by Iris Keren 博士の生徒たち: アリソン・アルメイダ、マシュー・コッツォリーノ、トゥルン・グエン、ジェシカ・ウォン
NEUF2011