- トリケラトプスなどケラトプス恐竜の起源
- Variation in Frills and Horns of the same species
- トリケラトプス・サーガ… 種の大集合。 16種から2種へ
- Triceratops vs. Torosaurus – Triceratops is not Torosaurs
- Horns and Frills – They Weren’t Made For Ramming!
- Triceratops Behavior.CERRATOPS OF THE DINOSAURS OF THE HEROONES: Herding Behavior
- Teeth and Diet
- How fast could Triceratops Dinosaur Run? トリケラトプスとセラトプシドのロコモーション
- Triceratops Dinosaur Skin – Scales, Scutes, and Quills??
- おすすめティラノサウルス本・化石:
トリケラトプスなどケラトプス恐竜の起源

CMNHにあるProtoceratops andrewsiの標本。
The Ceratopsids – Origins and Distribution
The Ceratopsidae Dinosaur familyは、顔の角と大きなフリルを特徴とする4脚の草食動物を包括しています。 1887年に発見されたトリケラトプスもこの仲間です。 セラトプス類には多くの亜科と属がある。 角やくちばし、フリルなどの形や大きさで分類されます。
ケラトプス類は、北米とアジアに限られています(トリケラトプスは北米西部のみ)。 原始的なケラトプス類はアジアで発見されており、アジアで最初に出現し、その後陸橋で北米に渡ったことがわかります。 白亜紀末に絶滅するまで、北米で急速に多様化した。 現在、セラトプス類は約37属に分類されている。

Ceratopsid Dinosaurのいくつかの属の頭蓋骨の画像です。 フリル、くちばし、角のワイルドなバリエーションに注目してください。
By skinnylawyer from Los Angeles, California, USA , via Wikimedia Commons
Variation in Frills and Horns of the same species
1900年代初頭にケラトプス類が見つかったとき、断片化された遺物はフリル、くちばし、角などの形で区分けされていました。 幼体か成体か、また性的二型性については考慮されませんでした。 その後、より多くの化石が発見されるにつれ、古生物学者は、動物が異なる発達段階に入ると頭蓋骨の形が変化することに気づいた。 角やフリルは大人になるまで完全に発達しませんでした(Farlow 327)
この素晴らしい例は、2013年に発見されたカスモサウルス科のセラトプシス恐竜の赤ちゃんからです。 2016年にCurrieらは、この赤ちゃん恐竜に関する論文を発表し、成体のカスモサウルス標本と比較しました(Currie et al, 2016)。 赤ちゃんカスモサウルスのフリルは、成体のフリルと全く違うように見えます。
トリケラトプス・サーガ… 種の大集合。 16種から2種へ

シカゴのフィールド自然史博物館に展示されているトリケラトプス恐竜の骨格
セラトプス類の数を減らす良い例がトリケラトプス属です。 トリケラトプスは、アメリカ西部のヘル・クリーク層で最も多く発見されている恐竜です。 トリケラトプス属はかつて、発見されたさまざまな頭骨のバリエーションから、最大で16種が割り当てられていました。 しかし、古生物学者がトリケラトプスのフリルの形が変化していることに気づき、再分析されることになりました。 1986年、オストロムとウェルンホーファーはトリケラトプス属を1種に絞り込み、T. horridusとした。 その後、1996年にキャサリン・フォレスターがトリケラトプスの種を調査し、T. horridus が T. prorsus よりも数が多いので、2つの種は異なる種であると結論づけました。 そして2009年、スカネラ氏とファウラー氏がこの見解を支持し、ついにこのサガは終わったのです。 トリケラトプスは2種あった…みたいな。 トロサウルスの問題もあるし…
Triceratops vs. Torosaurus – Triceratops is not Torosaurs

Torosaurus latus標本の大きさを比較したところ。
“A, YPM 1831, サブアダルト、頭骨長 >2.6 m, B, ANSP 15192, 初期アダルト、頭骨長 1.8 m。” YPM 1831の吻は復元されており、おそらくANSP 15192と同様に若干長かったと思われます。 からです。 Nicholas R. Longrich*, Daniel J. Field, 2012 – PLOS one under CC2 license.
トロサウルスはトリケラトプスと同じ地層で見つかった大きなケラトプスの仲間です。 より大きく、フリルも長く、上部付近に大きな穴が開いているのが特徴的です。 2010年、スカネラ氏とホーナー氏は、トロサウルスが実はトリケラトプスの後期急成長段階であることを証明する論文を発表した。 これが、さまざまな論争を巻き起こした。 古生物学者たちは、ある者は賛成し、ある者は反対したのです。 個人的には、顎骨に2つの大きな穴が単純に浸食されていることにいつも疑問を感じていました。 そうすると、筋肉の付着構造全体が一変してしまうのです。 ここ数年、スカンネラとホーナーの結果を否定する論文が多く発表されている。 2011年には、恐竜の専門家であるアンディ・ファーク博士が、トロサウルスは「トリケラトプスやネドケラトプスとは異なる属である」とする論文を発表しています。 最近の論文では、2012年のロングリッチとフィールドによる論文など、別属を支持する傾向がある。 彼らは様々なトロサウルスとトリケラトプスの頭蓋骨を研究し、それらを成長系列に並べるクラスタリング分析を行った。 彼らは、トロサウルスとトリケラトプスの両方に、未熟な個体と成熟した個体を発見したのです。 したがって、これらは同じ動物である可能性があります。 トリケラトプスとトロサウルスは別の恐竜である
上の画像はLongrich and Field (2012)より。
Horns and Frills – They Weren’t Made For Ramming!

Ceratopsid Dinosaur skull from Utah showing the elaborate Horns and Frills on display at the CMNH
Diceratops と Triceratops を除いて、ほとんどのセラトプス類には大きなフリル (鰓蓋) があります。 この2つの属は比較的縮小されたフリルをもっていた(Fastovsky p.180)。 生前のフリルの大きさは、顎の筋力に直接影響する(フリルが大きいほど、顎の筋力は大きい/強い)(Fastovsky p.186)。
角については、トリケラトプスなどの恐竜の化石を見ると忘れてしまう人が多いのですが、頭蓋骨にある角は骨芯です。 この骨芯は実際の角で覆われていたはずで、生前はもっと長かったはずです。
残念ながら、角やフリルが実際に何に使われていたかを確認することはできません。 しかし、現代の群れで暮らす動物に例えると、支配や防御、見栄えのために使われていたような気がします。 セラトプス類は、縄張り意識や交尾の権利のために、同種内で角を使っていたのかもしれない。 これは化石の証拠と一致します。トリケラトプスの頭蓋骨やフリルには、他のトリケラトプスから刺された傷がよく見つかり、何らかの種内戦闘があったことを示しています (Farke, et al, 2009)。 しかし、セラトプシドの角は、哺乳類の角とは異なる形をしていました。 ぶつかるのではなく、単に角を突き合わせて戦っていたのかもしれない。 模型を使った研究で、トリケラトプスが角をロックする行動をとることは物理的に可能であることがわかった(Farke, Andrew, A. 2004)。 彼らは3種類の角のロックが可能であることを発見した。 これは、ヘラジカが戦うときに角をロックするのに似ています。
興味深いことに、1つの例外があります。 セントロサウルスという大きな鼻の角が1本しかないセラトプス類は、ファーク(Farke, et al, 2009)によって研究され、傷がないことが判明しました。 このことは、セントロサウルスが角やフリルを交尾や縄張り争いに使わず、孔雀のようにディスプレイのために使っていたことを意味している。 ファーケの論文からの図を下に示す。 7089>
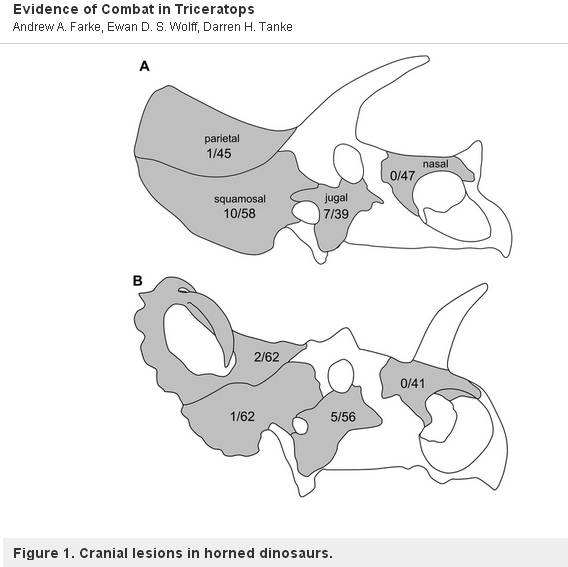
「(A)トリケラトプスと(B)セントロサウルスの頭蓋骨の模式図で、各頭蓋要素における病変(骨膜反応骨と骨折胼胝)の発生率(異常要素数/総要素数)を示す。 ノンスケール」
より。 Andrew A. Farke, Ewan D. S. Wolff, Darren H. Tanke, 2009
– PLOS one under CC2 license.
Moral of the story, Triceratops used them for mating behavior and Centrosaurus used them for display, 角とフリルはおそらく異なるタイプのセラトプス類によってさまざまな方法で使用されていたのだと思われます。
多くの角やフリルの用途の1つは、おそらく捕食者を追い払うためだったと思われます。 T-レックスに噛まれた跡が残っているものもあり、これはT-レックスの攻撃を食い止めることに成功した標本です。 一方、T-REXに噛まれた痕が治癒していないものも多く発見されており、これはT-REXの餌であったことを意味する。 このような化石の発見から、白亜紀後期のアメリカ西部に生息していた強大なT-レックスを撃退するために、フリルや角も使われたと考えられなくもない。
角やフリルは、角をロックするため、頭を束ねるため、ディスプレイ、防衛、または上記のすべてのために使われたかどうかはわからないが、ひとつだけ確かなことは、このフリルや角は、T-レックスを撃退するために使われたのだということだ。 下の画像を見てください!

Ceratopsid Dinosaurs horns and frills on display at the Carnegie Museum of Natural History.notice the first skull to the left (Pachyrhinosaurus sp.) has no horn at all, instead it has a thick bone knob.
Triceratops Behavior.CERRATOPS OF THE DINOSAURS OF THE HEROONES:
Herding Behavior
Centrosaur 100の化石の大きな骨層は中西部で発見されており、彼らが大きな群れで移動していたことを示している。 このことは、鹿、ガゼル、水牛のような群れをなす動物に角がよく使われていることと一致している。

Hell Creek Dinosaur Dig でサウスダコタ州の Hell Creek Formation から見つかった 3 つのトリケラトプスの歯の画像
Teeth and Diet
他の群れ行動と同様、セラトプスは草食性動物でした。 彼らの口は、かたい植物を食べるのに理想的な形をしていました。 これらの恐竜は、歯根が2本あり、歯列弓と呼ばれるグループに配置されていました。 歯列弓とは、隣接する歯が縦列と横列に並んで固定されているユニークな歯並びのことである。 各列には少なくとも3本の歯があり、1本は機能的で、2本は “代用品 “でした。 恐竜の生涯を通じて交換され続けた歯は、非常に垂直に配置され、ハサミのように剪断できるほど鋭かった(実際には自己研摩)。 咀嚼の際、この恐竜の歯は本当に人間のように全く触れず、その代わりに、切り裂くような動きで、互いの間をすり抜けた(ファストフスキー183)。 この鋭い歯と非常に強い顎の筋肉から、彼らはしばしば「自然界初の料理人!」と呼ばれる。 (Farlow 85)と呼ばれている。
余談だが、他の一般的な草食動物であるハドロサウルスも、同様の歯冠を備えていた。 彼らは、比較的低い草丈のため、被子植物、シダ類、その高さに生える小さな針葉樹などの低成長植物を食べていたのではないかと考えている。 その大きく重い頭蓋骨のために、後ろ足で立ち上がることはできなかったが、アメリカ西部の芝刈り機は、木の葉に近づくためにブルドーザーで木を乗り越えることができたかもしれない。

私が行ったヘルクリーク恐竜発掘で、ヘルクリーク層にまだ埋まっているトリケラトプスの歯の画像。
How fast could Triceratops Dinosaur Run? トリケラトプスとセラトプシドのロコモーション
多くの人がトリケラトプスとは非常にゆっくりでのんびりした動物だと思い込んでいます。 多くの博物館にあるセラトプス座恐竜の骨格復元(この記事の写真のものを含む)は、トリケラトプスがトカゲのようなのびのびとした姿勢をしていることを示しています。 トカゲのような姿勢は、セラトプス類の肋骨や脊椎骨を取り付ける際の誤差からきています。
上記の発言を裏付ける証拠は、トリケラトプスや他のセラトプス類の痕跡の分析から得られました。 ポールとクリスチャンセンは、2000年に発表した論文で、トラックウェイのペダルの印象を研究し、手と足の配置と角度を分析したのである。 この情報から、彼らはセラトプス類が象のような前肢の姿勢をしていることを突き止めた。 哺乳類のような姿勢によって、セラトプス類は爬虫類のような姿勢よりもはるかに速く走ることができるのです。 ポールとクリスチャンセンは、最も大きなケラトプス類の最大走行速度を “サイのそれとほぼ同じ “であると示した。 ちなみに、クロサイは時速55キロ、つまり34マイル(約15キロ)で走ることができるそうです

カーネギー自然史博物館のトリケラトプスの骨格復元では、トカゲのようなのびのびとした姿勢を見せていますが、これは最近の証拠では正しくありません。

シカゴのフィールド博物館からのトリケラトプス・モデルは、最近の証拠ではより哺乳類のように正しい姿勢を示します。
Triceratops Dinosaur Skin – Scales, Scutes, and Quills??
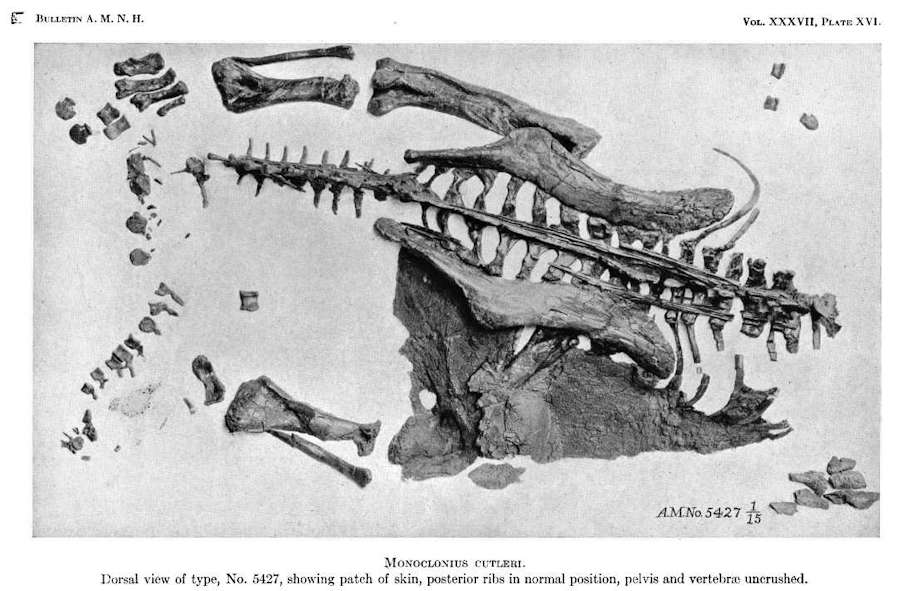
Image of the Centrosaurus dinosaur specimen (AMNH 5427) complete with a large skin patch
年代:1917、著者:Marcus, Inc: Barnum Brown,出典: AMNDHデジタルライブラリー。 B037a10.pdf Public Domain.
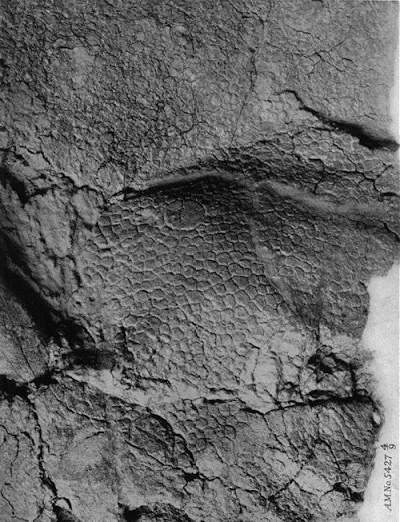
Closeup of the AMNH 5427 Centrosaurus dinsoaur skin impression.
Date: 1917, Author: Barnum Brown,出典: B037a10.pdf Public Domain.
恐竜の皮膚痕の化石は、非常に珍しいです。 皮膚付きの恐竜の化石を「ディノマミー」と呼ぶことがありますが、これはミイラ化したものではありません。
幸い、トリケラトプスを愛する人たちのために、「セラトプス類のミイラ」がいくつか発見されています。 上の写真のAMNH 5427は、皮膚の大きなパッチが保存されているセントロサウルスです。
より最近の「トリケラトプスのミイラ」は、2002年にワイオミング州でレーンという名で発見されました。 ブラックヒルズ地質研究所が慎重に、そして丹念に発掘したT. horridusの標本である。 この標本は現在、ケルシーと呼ばれるほぼ無傷のホリダスの骨格標本とともに、同博物館で展示されている。
トリケラトプスのレーンは、発見された恐竜の化石の中で、最も皮膚の保存状態が良い。 数フィートの皮膚が保存されている箇所もある。
その皮膚は、多くの古生物学者が考えていたものとは全く異なっています。 下面は、ワニに似たスクートで覆われている。 恐竜の他の部分は、小さな鱗で覆われています。 しかし、不規則な鱗のような形もある。 こぶし型の六角形で、中央に小さな孔のような穴が開いているものもある。
この穴から羽毛が飛び出していたのではないかと推測する人もいる。 この「羽ペン」仮説は、トリケラトプスの親戚で、アジアに生息するプシッタコサウルスに由来しています。 これは遠縁の親戚で、尾の先端に羽のような構造を持っている。 ただし、プシッタコサウルスはトリケラトプスの遠い親戚なので、トリケラトプスにも羽毛があったというわけではないことに注意が必要である。 羽毛」仮説は現在のところ推測の域を出ておらず、トリケラトプスの皮膚の正確な性質については、今後の研究・出版を待つ必要がある。

六角形の鱗が見える恐竜レーンのトリケラトプス皮膚キャストの断面。 Image by:Black Hills Institute of Geological Research

保存された羽毛と模型を持つピタコサウルス類。 古生物学者は、この羽の構造はトリケラトプスの奇妙な鱗の一部から突き出ているのではないかと推測しています。
おすすめティラノサウルス本・化石:

New Perspectives on Horned Dinosaurs: The Royal Tyrrell Museum Ceratopsian Symposium (Life of the Past)
Ceratopsianについて最新見解を示す本書は、ティラノサウルスの角竜について解説しています。 角竜の機能、生物学、行動、古生態、古地理に関する新しい研究を紹介しています。 また、多くの新種についても記述しています。 学術論文集なので、本格的な恐竜読者限定!? 恐竜好きにはたまらない参考書だと思います。

Dinosaurs Rediscovered: 古生物学の科学革命
著 マイケル・J・ベントン 2019年
恐竜の一般書として最適です。 最新の内容で、恐竜に対する私たちの見方を完全に変えた過去20年間の新発見について解説しています。 色、スピード、噛む力、子供の世話など…ほんの数年前までは知らなかったことを簡単に章立てしています!レビューやサンプルページをご覧ください!

High quality Dinosaur teeth by Fossilera
References / Works Cited
Currie Philip J., ホームズ・ロバート・B.・J・ライアン・マイケル・コイ・クライブ (2016) カナダ・アルバータ州ダイナソーパーク層から出土したカスモサウルス型セラトプシド(恐竜界・鳥脚類)幼生。 Journal of Vertebrate Paleontology, 2016; e1048348 DOI: 10.1080/02724634.2015.1048348
Farlow, O. James, Brett-Surman, M.K., Editors (1997). ザ・コンプリート・ダイナソー. IN: Indiana University Press.
Fastovsky, E. David, Weishampel, B. David (1996). 恐竜の進化と絶滅. NY: 7838>Farke A.A. (2011) Anatomy and Taxonomic Status of the Chasmosaurine Ceratopsid Nedoceratops hatcheri from the Upper Cretaceous Lance Formation of Wyoming, U.S.A. PLoS ONE 6(1): e16196. doi: 10.1371/journal.pone.0016196
Farke A.A., Wolff EDS, Tanke DH. (2009)トリケラトプスにおける戦闘の証拠。 PLoS ONE 4(1): e4252. doi: 10.1371/journal.pone.0004252
Farke, A.A. 2004年. トリケラトプス(Dinosauria: Ceratopsidae)の角の使用。 スケールモデルによる行動仮説の検証。 古生物学エレクトロニカ7(1):10 pp. (PDF)
Forster Catherine A. (1996) トリケラトプスにおける種の解明:系統的・形態的アプローチ. Journal of Vertebrate Paleontology, 16:259-270.
Longrich・NR, Field DJ. (2012) Torosaurus Is Not Triceratops: (2012) Torosaurus Is Not Triceratops: Ontogeny in Chasmosaurine Ceratopsids as a Case Study in Dinosaur Taxonomy. PLoS ONE 7(2): e32623. doi:10.1371/journal.pone.0032623
Ostrom J.H., Wellnhofer P. (1986) The Munich specimen of Triceratops with a revision of the genus.ミュンヘンのトリケラトプス標本とトリケラトプス属の修正. Zitteliana. 14, 111-158.
Scannella J, Horner JR. (2010) Torosaurus is Triceratops, synonymy through ontogeny. Journal of Vertebrate Paleontology 30: 1157-1168.
Scannella, J.B. and D.W. Fowler.邦訳は「脊椎動物古生物学」. (2009)トリケラトプスにおけるアナジェネシス:ヘル・クリーク層の新しく解決された層序フレームワークからの証拠. 北アメリカ古生物学会議 , アブストラクト集: 148-9.
Gregory S. Paul, Per Christiansen. (2000)新ケラトプス類の恐竜における前肢の姿勢:歩行と運動への示唆 古生物学サマー,v.26 no.3 p. 450-465
.